


Pompa sodio-potassio
TOPICS
Definizione

La verifica dello stato stazionario delle membrane cellulari, condotta applicando l’equilibrio elettrochimico di Nernst a ciascun ione, evidenzia che la distribuzione ionica osservata in molte cellule non può essere spiegata unicamente dai meccanismi di diffusione passiva. Occorre quindi invocare un trasporto dipendente da processi biochimici localizzati in proteine di membrana: si tratta del trasporto attivo. Nel caso in esame, l’elemento chiave è la pompa sodio‑potassio (Na⁺/K⁺), capace di spostare Na⁺ verso l’esterno e K⁺ verso l’interno contro i relativi gradienti elettrochimici, utilizzando energia metabolica.
Numerose evidenze sperimentali e considerazioni fisiche portano a postulare un dispositivo di trasporto attivo che estrude Na⁺ e, in contropartita, introduce K⁺. In questa sede non si entra nel dettaglio molecolare del ciclo catalitico, ma si mettono in relazione, con argomentazioni fisiche, il contributo della pompa sodio‑potassio e i flussi passivi con la distribuzione ionica e il potenziale di membrana mostrati in (Figura 06.14-01) e (Figura 06.14-02).
Principali evidenze a favore di una pompa Na⁺/K⁺ attiva:
- le concentrazioni intracellulari ed extracellulari di Na⁺ si discostano da quelle previste dal solo equilibrio di Nernst; analogamente, le concentrazioni di K⁺ non corrispondono esattamente al valore di equilibrio teorico;
- pur essendo la membrana parzialmente permeabile al Na⁺ (Pᴺᵃ ≠ 0), le concentrazioni di Na⁺ restano sostanzialmente stabili nel tempo, benché risultino opposte a quelle di equilibrio di Nernst;
- l’efflusso di Na⁺ decresce marcamente quando si inibisce il metabolismo energetico cellulare o si abbassa la temperatura, indicando dipendenza da un processo endoergonico;
- il flusso di Na⁺ è accoppiato a quello di K⁺: riducendo la concentrazione extracellulare di K⁺ diminuisce l’estrusione di Na⁺, mentre l’aumento di K⁺ esterno facilita l’efflusso di Na⁺.
Un modo essenziale per formalizzare l’ipotesi della pompa sodio‑potassio, considerata ione per ione, è imporre che la somma dei flussi totali (passivi + attivi) di Na⁺ e K⁺ sia nulla, condizione tipica dello stato stazionario:
\[J_{\text{Na}} (\text{totale}) + J_{\text{K}} (\text{totale}) = 0,\]
da cui, risolvendo rispetto al potenziale di membrana Vₘ, si ricava:
\[V_m = \frac{RT}{ZF} \ln \left[\frac{\{\text{K}^+\}_e + \frac{P_{\text{Na}}}{P_{\text{K}}}\{\text{Na}^+\}_e}{\{\text{K}^+\}_i + \frac{P_{\text{Na}}}{P_{\text{K}}}\{\text{Na}^+\}_i}\right],\]
nota come equazione di Goldman.
Nota la composizione ionica, la (V_m = \frac{RT}{ZF} \ln \left[ \frac{\{\text{K}^+\}_e + \frac{P_{\text{Na}}}{P_{\text{K}}} \{\text{Na}^+\}_e}{\{\text{K}^+\}_i + \frac{P_{\text{Na}}}{P_{\text{K}}} \{\text{Na}^+\}_i} \right]) consente di ricavare il rapporto delle permeabilità se Vₘ è misurato, e viceversa. Per cellule eccitabili a riposo si ottiene tipicamente Pᴺᵃ/Pᴷ ≈ 10⁻2, in accordo con la P_{\text{Na}} = 10^{-8} \text{ cm/s} \quad \text{e} \quad P_{\text{K}} = 10^{-6} \text{ cm/s}. Dalla (V_m = \frac{RT}{ZF} \ln \left[ \frac{\{\text{K}^+\}_e + \frac{P_{\text{Na}}}{P_{\text{K}}} \{\text{Na}^+\}_e}{\{\text{K}^+\}_i + \frac{P_{\text{Na}}}{P_{\text{K}}} \{\text{Na}^+\}_i} \right]) si vede inoltre che, quando Pᴺᵃ/Pᴷ ≪ 1, Vₘ si approssima al potenziale di Nernst del potassio, Vₘ₍ₖ₎, come osservato sperimentalmente. Una conseguenza rilevante è l’elevata sensibilità di Vₘ a variazioni del rapporto Pᴺᵃ/Pᴷ: piccole modifiche di permeabilità al Na⁺ possono produrre scostamenti significativi del potenziale di membrana.
La condizione J_{\text{Na}}(\text{totale}) + J_{\text{K}}(\text{totale}) = 0 descrive uno stato stazionario dinamico, non un equilibrio termodinamico. I flussi passivi di “perdita” (leak) attraverso canali aperti vengono continuamente compensati dall’azione della pompa sodio‑potassio, che utilizza energia metabolica per ripristinare i gradienti. Ciò permette di mantenere un’elevata asimmetria di Na⁺ e K⁺ ai lati della membrana e, quindi, di sostenere il valore di Vₘ previsto dalla (V_m = \frac{RT}{ZF} \ln \left[ \frac{\{\text{K}^+\}_e + \frac{P_{\text{Na}}}{P_{\text{K}}} \{\text{Na}^+\}_e}{\{\text{K}^+\}_i + \frac{P_{\text{Na}}}{P_{\text{K}}} \{\text{Na}^+\}_i} \right]). Dal punto di vista energetico, la pompa è un sistema accoppiato: il lavoro chimico interno alla proteina di membrana alimenta il trasporto contro gradiente e rende possibili i successivi flussi passivi accoppiati.
È utile ricordare che la pompa Na⁺/K⁺ è elettrogenica: per ogni ciclo, in condizioni fisiologiche, vengono tipicamente espulsi 3 Na⁺ e introdotti 2 K⁺, con una perdita netta di cariche positive dall’interno verso l’esterno. Questo contribuisce direttamente a Vₘ con pochi millivolt (circa 2,5–5 mV in molte cellule), mentre la parte predominante del potenziale a riposo deriva dalla selettività di permeabilità, in particolare verso K⁺. Inibitori specifici della pompa riducono l’estrusione di Na⁺, provocano accumulo intracellulare di Na⁺, diminuzione di K⁺ e, nel tempo, depolarizzazione, confermando il ruolo della pompa nel mantenimento dello stato stazionario.
Il funzionamento complessivo della pompa sodio‑potassio e la sua interazione con i flussi passivi è rappresentato schematicamente in (Figura 06.14-03). Tale meccanismo giustifica l’equilibrio dinamico osservabile nella membrana delle fibre muscolari (Figura 06.14-03) e, in termini qualitativi analoghi, si applica anche ad altre cellule, come gli eritrociti menzionati alla fine del paragrafo precedente, dove la pompa contribuisce anche al controllo del volume cellulare contro il gonfiore osmotico.
Oltre alla Na⁺/K⁺, altre pompe e trasportatori attivi assicurano gli assetti ionici caratteristici di specifici epiteli e tessuti: ad esempio, nelle membrane dei tubuli renali operano pompe protoniche e scambiatori accoppiati a gradienti preesistenti, che modulano pH e riassorbimento elettrolitico. L’impostazione descrittiva resta analoga: uno stato stazionario nasce dall’interazione fra permeabilità selettive (trasporto passivo) e meccanismi che consumano energia metabolica per stabilire e mantenere differenze di concentrazione e di potenziale.
Image Gallery
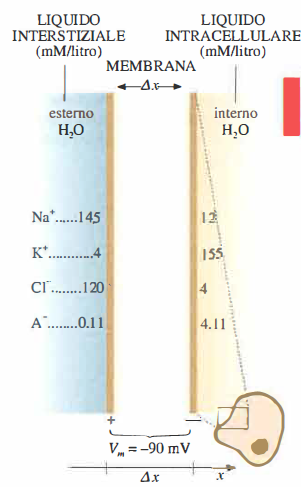
Image Gallery
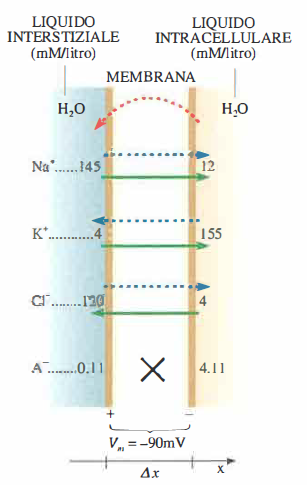
Image Gallery
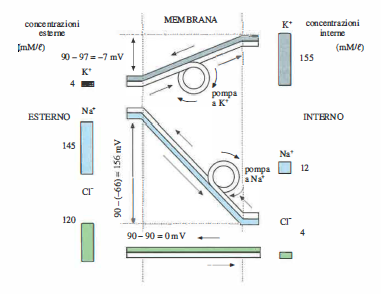
Image Gallery


