


Mandibola negli animali
Definizione

Le condizioni di equilibrio meccanico vincolano in modo sostanziale la forma e le dimensioni delle strutture biologiche, influenzando i percorsi di differenziazione evolutiva anche quando le strutture sono deformabili. Un caso esemplare è l’apparato masticatorio: l’analisi comparativa dell’articolazione mandibolare nei rettili e nei mammiferi, con un ulteriore confronto tra forme carnivore ed erbivore, mostra come la direzione, il punto di applicazione e l’intensità delle forze muscolari abbiano rimodellato l’architettura cranio-mandibolare. Poiché ossa, muscoli e articolazioni assolvono funzioni meccaniche precise, le loro proporzioni sono in larga misura dettate dai carichi funzionali, in particolare dalla necessità di generare una forza di morso elevata senza porre a rischio l’integrità dell’articolazione temporo-mandibolare.
I reperti paleontologici indicano che i mammiferi discendono da sinapsidi affini ai rettili, nei quali la mandibola inferiore era composta da più elementi e l’articolazione coinvolgeva le ossa quadrato e articolare. Nel corso dell’evoluzione dei mammiferi si è affermata l’articolazione dentario-squamoso, mentre parte degli elementi postdentari ha dato origine agli ossicini dell’orecchio medio; in parallelo, la superficie di inserzione dei muscoli elevatori della mandibola si è ampliata (sviluppo del processo coronoideo), e la direzione delle risultanti muscolari si è riorientata, con importanti conseguenze sull’equilibrio dell’articolazione e sulla sicurezza del morso. Ciò consente, a parità di forza trasmessa al cibo, di ridurre le sollecitazioni sul fulcro articolare.
Nella (Figura 02.21-01) sono schematizzate due configurazioni tipiche della mandibola inferiore: nei rettili (a) la mandibola si comporta come una barra con il muscolo principale che esercita una forza Fm diretta superiormente e applicata vicino al fulcro F; nei mammiferi (b) è presente un pronunciato processo coronoideo per l’inserzione del muscolo temporale, che esercita una forza Fm1 diretta posteriormente e in alto, mentre i muscoli massetere e pterigoideo imprimono una forza Fm2 con componente anteriore e superiore. Consideriamo come tali configurazioni influenzino le condizioni di equilibrio e le sollecitazioni sull’articolazione durante il morso.
Per un rettile primitivo (Figura 02.21-01 ), quando sull’alimento si applica una forza di presa −Fr con i denti posteriori, la mandibola riceve una reazione +Fr diretta inferiormente. L’equilibrio traslazionale richiede l’intervento della reazione vincolare R al fulcro, che controbilancia la risultante delle forze:
\[
\mathbf{F}_m = \mathbf{R} + \mathbf{F}_r, \quad \text{cioè} \quad \mathbf{F}_m = \mathbf{R} + \mathbf{F}_r,
\]
Poiché il punto di applicazione di Fm è vicino al fulcro, il bilancio dei momenti rispetto a F porta a:
\[
x_{F_r} F_r - x_R R = 0, \quad \text{da cui} \quad R = \frac{x_{F_r} F_r}{x_R}.
\]
segue quindi che la forza muscolare complessiva necessaria è:
\[ F_m = R + F_r = F_r \left( 1 + \frac{x_{F_r}}{x_R} \right) \]
La mandibola del rettile si comporta quindi da leva di terzo genere, con guadagno meccanico G inferiore all’unità (leva svantaggiosa):
\[
G = \frac{x_R}{x_R + x_{F_r}}
\]
Ne deriva che, a parità di forza sul cibo Fr, sia la reazione articolare R sia lo sforzo muscolare risultano maggiori; di conseguenza il carico sul fulcro pone un vincolo superiore alla forza di morso utilizzabile senza rischio di frattura o lussazione.
Esempio alternativo: se xF_r = 3 xR e Fr = 1,2 N, allora R = 3,6 N e Fn = 4,8 N, con guadagno G = 1/(1+3) = 0,25. La forza effettiva sul bolo alimentare è, dunque, nettamente inferiore rispetto alle forze muscolare e di reazione articolare.
Nei mammiferi, la forza Fm1 agisce a un braccio più lungo e si aggiunge la risultante Fm2 di massetere e pterigoideo (Figura 02.21-01 ). Quando le linee d’azione di Fm1, Fm2 e Fr sono concorrenti, i loro momenti rispetto al punto d’incontro sono nulli; l’equilibrio dei momenti impone che anche la reazione vincolare R sia applicata lungo la stessa direzione. Se inoltre la somma vettoriale delle tre forze è nulla, si ha:
\[
\mathbf{F}_{m1} + \mathbf{F}_{m2} + \mathbf{F}_r = 0
\]
in questo caso non è necessaria alcuna reazione vincolare R per l’equilibrio traslazionale. Anche quando la condizione non è soddisfatta esattamente o le linee non sono perfettamente concorrenti, la reazione R resta molto inferiore rispetto al caso dei rettili. Si comprende così perché l’articolazione dei mammiferi possa essere più compatta senza compromettere la sicurezza meccanica, permettendo al contempo un più ampio sviluppo della muscolatura masticatoria.
Considerando l’equilibrio dei momenti soddisfatto e ponendo R = 0, dalla (Figura 02.21-02) si ricavano con maggiore dettaglio le condizioni d’equilibrio traslazionale in componenti:
\[
\begin{cases}
F_{m1} \cos \theta_1 - F_{m2} \cos \theta_2 + F_r \cos \theta_r = 0 \\
F_{m1} \sin \theta_1 + F_{m2} \sin \theta_2 - F_r \sin \theta_r = 0
\end{cases}
\]
Assumendo, per semplicità, θ1 = θ2 = 45° e indicando con Fm2 = k Fm1 il rapporto tra la forza dei muscoli massetere-pterigoideo e quella del temporale, diventano:
\[
\begin{cases}
F_r \cos \theta_r = F_{m1} \dfrac{k - 1}{\sqrt{2}} \\
F_r \sin \theta_r = F_{m1} \dfrac{k + 1}{\sqrt{2}}
\end{cases}
\]
da cui si ricavano:
\[
\tan \theta_r = \frac{k + 1}{k - 1}
\quad \text{e} \quad
F_r = F_{m1} \sqrt{k^2 + 1}
\]
Queste relazioni consentono di discutere differenze funzionali tra carnivori ed erbivori. Nei primi, i denti anteriori e i premolari sono impiegati per afferrare e lacerare, con una grande partecipazione del muscolo temporale; negli erbivori prevale la triturazione occlusale tra molari, con potenziamento relativo del sistema massetere-pterigoideo e un apparato articolare che favorisce ampie escursioni latero-protrusive.
Per i carnivori si assume spesso che la massa del temporale rappresenti circa 2/3 della muscolatura adductoria totale, mentre negli erbivori il temporale costituisce una frazione più ridotta, per esempio 1/10. Ne segue, rispettivamente, k = 0,5 e k = 9, con i risultati:
\[ \operatorname{tg}\theta_r = \frac{1.5}{-0.5} = -3.0, \]
quindi:
\[
\theta_r = 108.4^\circ \quad \text{e} \quad F_r \approx 1.1 F_{m1}
\]
ossia la risultante sul cibo è orientata anteriormente e ha modulo confrontabile con la forza del temporale (Figura 02.21-03). Per gli erbivori, con k = 9, si ottiene:
\[ \operatorname{tg}\theta_r = \frac{10}{8} = 1.25, \]
e dunque:
\[
\theta_r = 51.3^\circ \quad \text{e} \quad F_r \approx 9 F_{m1}
\]
vale a dire che la risultante è orientata posteriormente, a vantaggio della macinazione tra le superfici occlusali (Figura 02.21-03), e il suo modulo supera di molto la forza del temporale. In entrambi i casi, le pressioni sul fulcro restano contenute grazie all’assetto vettoriale delle forze, permettendo un rimodellamento evolutivo che dissocia la robustezza dell’articolazione dallo sviluppo delle masse muscolari:
- Determinanti meccanici della forza di morso: bracci di leva dei muscoli elevatori; angoli d’inserzione e direzione delle linee d’azione; posizione del punto di contatto sul dente rispetto al fulcro;
- Architettura muscolare: nei carnivori domina il temporale con processo coronoideo elevato; negli erbivori si espande la regione zigomatica per l’ancoraggio del massetere, con condilo mandibolare adattato a movimenti trasversali;
- Conseguenze articolari: nei mammiferi, la quasi-concorrenza delle forze riduce la necessità della reazione vincolare, consentendo articolazioni più leggere e resistenti alla fatica.
Esempio aggiuntivo comparativo: assumendo angoli come sopra e k = 0,7 (forma tendenzialmente carnivora), si ha \(\tan \theta_r = \frac{1,7}{-0,3} \approx -5,67\), quindi \(\theta_r \approx 100,0^\circ\) e \(F_r \approx 1,22\,F_{m1}\); per un erbivoro moderato con k = 6, \(\tan \theta_r = \frac{7}{5} = 1,4\), \(\theta_r \approx 54,5^\circ\) e \(F_r \approx 6,08\,F_{m1}\). Anche tali valori mostrano lo spostamento dell’orientazione della risultante e l’incremento relativo del contributo masseterico nelle forme erbivore.
Image Gallery
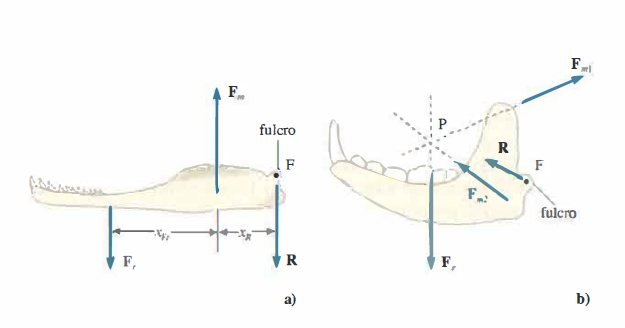
Image Gallery
Image Gallery
Image Gallery
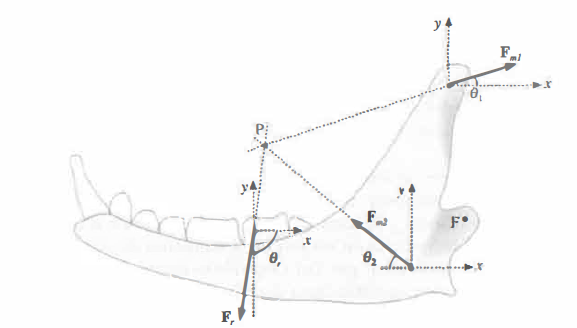
Image Gallery
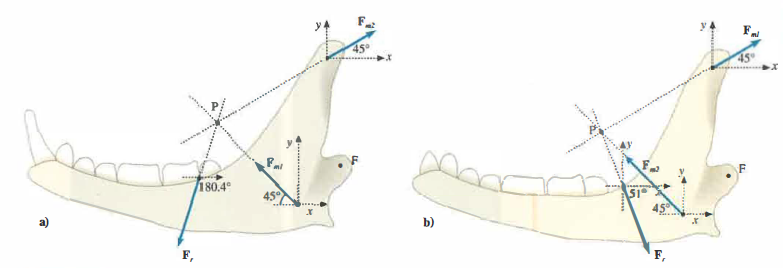
Image Gallery


