Le proprietà di un elemento dipendono dalla struttura degli atomi da cui è costituito
TOPICS
Definizione

Un atomo è organizzato attorno a un nucleo centrale compatto, positivamente carico, circondato da una distribuzione di elettroni negativi che occupano regioni di spazio quantistiche e sono mantenuti in prossimità del nucleo dall’attrazione coulombiana (Figura 01.08-01). Il nucleo ospita due specie di particelle subatomiche: protoni, dotati di carica positiva, e neutroni, elettricamente neutri. Il conteggio dei protoni, indicato con il numero atomico \(Z\), identifica l’elemento chimico. Così, l’idrogeno ha \(Z=1\) perché possiede un solo protone, mentre il carbonio ha \(Z=6\) in quanto contiene sei protoni (Figura 01.08-02). In un atomo isolato e complessivamente neutro, il numero di elettroni eguaglia il numero di protoni, perciò anche la popolazione elettronica è pari a \(Z\). Come noto, le proprietà chimiche di un elemento dipendono primariamente da \(Z\) e dalla configurazione degli elettroni più esterni.
I neutroni hanno massa pressoché equivalente a quella dei protoni e contribuiscono alla stabilità del nucleo, senza alterare direttamente il comportamento chimico. La variazione nel numero di neutroni genera isotopi di uno stesso elemento: specie con uguale \(Z\) ma diverso numero di neutroni \(N\). È utile introdurre anche il numero di massa \(A\), dato da \(A=Z+N\), e la notazione dei nuclei \(^{A}_{Z}\mathrm{X}\). Molti elementi presentano più isotopi naturali, alcuni dei quali sono instabili e vanno incontro a decadimento radioattivo. Il carbonio, per esempio, si trova prevalentemente come \(^{12}\mathrm{C}\) (sei protoni e sei neutroni), ma esiste anche l’isotopo \(^{14}\mathrm{C}\), con sei protoni e otto neutroni, che è radioattivo. Il suo decadimento, lento e con velocità pressoché costante nel tempo, è sfruttato per la datazione di materiali organici antichi, come documenti e resti biologici, grazie a un’emivita di circa 5.730 anni.
La massa atomica relativa di un atomo e la massa molecolare relativa di una molecola esprimono la massa rispetto a 1/12 della massa del carbonio-12. L’unità di misura è l’unità di massa atomica unificata, indicata come u, equivalente al dalton (Da): \(1\,\mathrm{u} = 1\,\mathrm{Da} = \tfrac{1}{12}\) della massa di \(^{12}\mathrm{C}\). Di conseguenza, per definizione, \(^{12}\mathrm{C}\) ha massa atomica 12, mentre \(^{14}\mathrm{C}\) ha massa atomica 14. Poiché gli elettroni sono circa 1/1836 della massa di un protone, il loro contributo alla massa totale è trascurabile nella maggior parte dei calcoli; un protone o un neutrone hanno massa di circa \(1\,\mathrm{Da}\), cioè approssimativamente \(1{,}67\times 10^{-24}\,\mathrm{g}\).
Le dimensioni atomiche sono estremamente ridotte: i raggi atomici tipici sono dell’ordine di \(0{,}1\text{–}0{,}3\,\mathrm{nm}\). Un atomo di carbonio ha un diametro di circa \(0{,}2\,\mathrm{nm}\); disporre una sequenza di atomi su una linea di \(1\,\mathrm{mm}\) richiederebbe dell’ordine di milioni di atomi, a sottolineare la scala microscopica della materia.
Il passaggio dal mondo microscopico a quello macroscopico è reso quantitativamente possibile dal numero di Avogadro, \(N_A\), che vale \(6{,}022\times 10^{23}\,\mathrm{mol^{-1}}\) (spesso approssimato come \(6\times 10^{23}\)). Per una sostanza di massa molecolare relativa \(M\), una mole corrisponde a \(N_A\) unità elementari (atomi, molecole o unità formula) e ha massa \(M\,\mathrm{g}\). La relazione fondamentale è: \[ n=\frac{N}{N_A},\qquad m = n\,M, \] dove \(N\) è il numero di particelle, \(n\) le moli e \(m\) la massa in grammi. La (Figura 01.08-03) illustra questa corrispondenza di scala:
- per l’acqua, \(M(\mathrm{H_2O})=18\); \(18\,\mathrm{g}\) contengono \(N_A\) molecole, mentre \(9{,}0\,\mathrm{g}\) ne contengono \(0{,}5\,N_A\);
- per il cloruro di sodio, \(M(\mathrm{NaCl})\approx 58{,}5\); \(58{,}5\,\mathrm{g}\) contengono \(N_A\) unità formula, e \(5{,}85\,\mathrm{g}\) corrispondono a \(0{,}10\,\mathrm{mol}\);
- per il diossido di carbonio, \(M(\mathrm{CO_2})=44\); \(44\,\mathrm{g}\) di \(\mathrm{CO_2}\) corrispondono a una mole di molecole e contengono \(2\,N_A\) atomi di ossigeno complessivi.
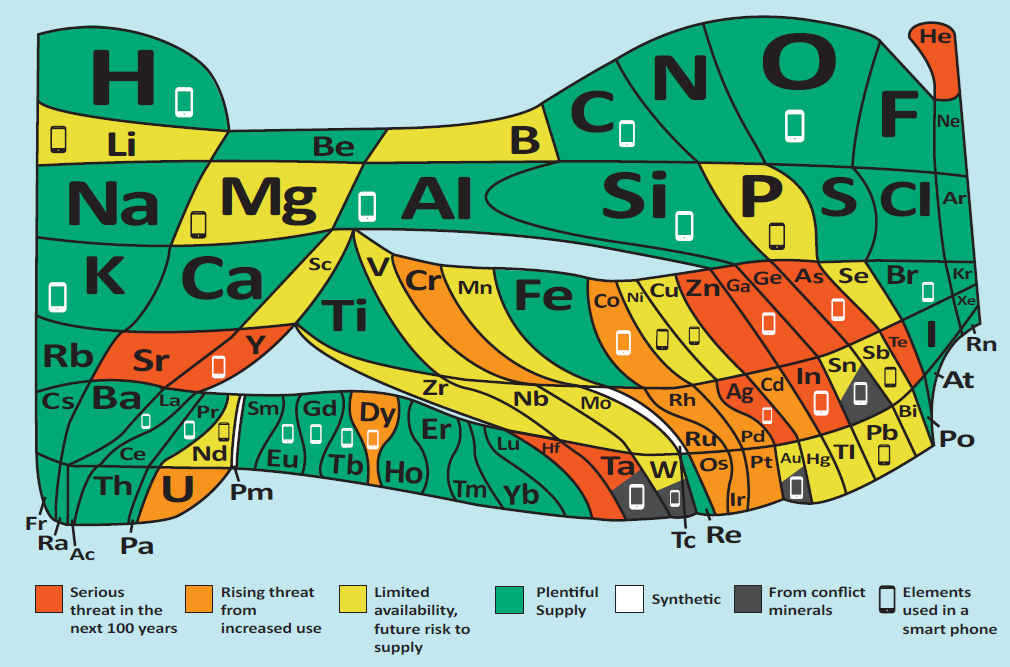
In natura sono presenti all’incirca 90 elementi, distinti per il valore di \(Z\) e per il numero di elettroni dei loro atomi neutri; tuttavia, la materia vivente è dominata da un set molto ristretto: carbonio (C), idrogeno (H), azoto (N) e ossigeno (O) costituiscono circa il 96% della massa degli organismi. Questa distribuzione è fortemente diversa da quella dei comparti inorganici del pianeta, come crosta terrestre e oceani (Figura 01.08-04), e riflette una chimica peculiare, centrata sulle proprietà di legame del carbonio e sull’onnipresenza dell’acqua. Altri elementi, pur quantitativamente minori, sono funzionalmente cruciali: fosforo e zolfo per acidi nucleici e proteine; sodio, potassio, magnesio, calcio e cloro per l’equilibrio ionico e la segnalazione; oligoelementi quali ferro, zinco e rame come cofattori enzimatici:
- numero atomico e neutralità: in un atomo neutro \(Z=\) protoni \(=\) elettroni;
- isotopia: isotopi di uno stesso elemento hanno uguale \(Z\) ma diverso \(N\); il numero di massa è \(A=Z+N\);
- unità di massa: \(1\,\mathrm{u}=1\,\mathrm{Da}=\tfrac{1}{12}\,\mathrm{massa}(^{12}\mathrm{C})\); la massa di protone e neutrone è circa \(1\,\mathrm{Da}\);
- scala mole: una quantità pari a \(N_A\) particelle ha massa \(M\,\mathrm{g}\) se \(M\) è la massa molecolare relativa; la mole collega conteggi atomici e misure in grammi.
In sintesi, benché il repertorio degli elementi sia ampio, la biochimica della vita sfrutta in modo preponderante pochi tipi di atomi, la cui combinazione e distribuzione elettronica determinano le reattività e le architetture molecolari alla base dei sistemi biologici.
Image Gallery

Image Gallery
.jpg)
Image Gallery

Image Gallery
Per comprendere come gli atomi si associno in molecole alla base dei sistemi viventi, è necessario concentrare l’attenzione sul comportamento degli elettroni. Proton i e neutroni restano confinati nel nucleo, stabilizzati dalle forze nucleari forti, e cambiano stato solo in condizioni fisiche estreme, come nel decadimento radioattivo, nel Sole o in un reattore nucleare. Nei sistemi biologici, invece, sono gli elettroni a ridistribuirsi tra atomi diversi: essendo la porzione più accessibile dell’atomo, determinano le regole della chimica e, quindi, le modalità con cui gli atomi si combinano in molecole.
Il moto elettronico su scala atomica non segue le leggi della meccanica classica ma quelle della meccanica quantistica. Gli elettroni occupano stati energetici discreti organizzati in gusci (strati) e sottogusci, descritti più correttamente in termini di orbitali. Ciascun guscio può ospitare un numero massimo di elettroni definito, e la loro occupazione è vincolata da principi quantistici (Aufbau, esclusione di Pauli e regola di Hund). In termini generali, la capacità massima del guscio di numero quantico principale \(n\) è data da \(2n^2\); tuttavia, negli elementi rilevanti per la biologia è utile una descrizione semplificata basata sull’ordine energetico degli orbitali più esterni.
Gli elettroni più vicini al nucleo, attratti con maggiore intensità dalla carica positiva nucleare, riempiono per primi il guscio interno, che contiene al massimo due elettroni. Il guscio successivo può ospitarne fino a otto; il terzo, per gli elementi leggeri comunemente presenti nelle biomolecole, è anch’esso saturabile con otto elettroni in condizioni tipiche; il quarto e il quinto strato, quando coinvolgono anche orbitali d, possono contenere fino a 18 elettroni. Atomi con più di quattro gusci occupati compaiono raramente nelle strutture molecolari biologiche. In generale, gli elettroni riempiono i livelli energetici dal più basso al più alto, e una configurazione è tanto più stabile quanto più gli elettroni occupano stati energetici profondi.
Un atomo che presenta il guscio di valenza completamente occupato è chimicamente poco reattivo. Esempi tipici sono i gas nobili: elio (2 elettroni, numero atomico 2), neon (2 + 8 elettroni, numero atomico 10) e argon (2 + 8 + 8 elettroni, numero atomico 18). Al contrario, l’idrogeno, con un unico elettrone e quindi un guscio incompleto, è altamente reattivo. Gli elementi prevalenti negli organismi (Figura 01.08-05) hanno tutti un guscio esterno non completo e possono quindi interagire per formare legami chimici. Poiché una configurazione con guscio pieno è energeticamente favorita, gli atomi con strato esterno incompleto tendono ad acquisire, perdere o condividere elettroni per completarlo. Le due modalità principali sono:
- trasferimento di elettroni da un atomo all’altro, con formazione di ioni e legami ionici;
- condivisione di coppie di elettroni tra due atomi, con formazione di legami covalenti (Figura 01.08-06).
L’idrogeno (H), cui manca un elettrone per saturare il primo guscio, tipicamente lo ottiene per condivisione, formando un legame covalente. Gli elementi più comuni nelle cellule, come carbonio (C), azoto (N) e ossigeno (O), con secondo guscio incompleto, e fosforo (P) e zolfo (S), con terzo guscio non saturo (Figura 01.08-05), tendono a condividere elettroni fino a raggiungere una configurazione di otto elettroni nel guscio esterno (regola dell’ottetto), spesso stabilendo più legami covalenti. Il numero di legami che un atomo può instaurare (valenza) è determinato da quanti elettroni deve acquisire o “mettere in comunione” per completare il proprio guscio di valenza.
La centralità degli elettroni più esterni nel definire le proprietà chimiche spiega la periodicità del comportamento degli elementi quando sono ordinati per numero atomico. Elementi con configurazioni di valenza analoghe mostrano reattività simile: ad esempio, un elemento con un unico elettrone nel secondo guscio e uno con un unico elettrone nel terzo presentano tendenze chimiche affini. In termini generali, i metalli possiedono pochi elettroni nel guscio esterno e li cedono con relativa facilità, mentre i gas nobili mostrano scarsa reattività poiché il guscio è già completo. Questa ricorrenza di proprietà è alla base dell’organizzazione della tavola periodica (Figura 01.08-07), in cui sono evidenziati anche gli elementi che costituiscono la materia vivente. Per chiarezza operativa, le valenze tipiche negli ambienti biologici sono spesso schematizzate come segue:
- H: 1 legame covalente;
- O: 2 legami covalenti;
- N: 3 legami covalenti;
- C: 4 legami covalenti;
- P: usualmente 5 legami nelle specie di fosfato; in alcuni contesti 3 legami;
- S: generalmente 2 legami; in composti specifici può raggiungere valenze maggiori.
Questa rappresentazione, seppure semplificata, è efficace per interpretare la formazione di molecole organiche e biomolecole complesse a partire da regole elettroniche elementari.
Image Gallery

Image Gallery

Image Gallery
Image Gallery

Le proprietà strutturali e funzionali delle cellule derivano dalle molecole che esse includono. Una molecola è un insieme di atomi uniti da legami covalenti, nei quali due atomi mettono in comune una o più coppie di elettroni, così da ottenere entrambi una configurazione elettronica più stabile. Nel caso più semplice, la molecola di idrogeno (H₂), ciascun atomo di H contribuisce con un elettrone e la coppia condivisa consente a entrambi di completare il primo guscio. La densità elettronica condivisa si concentra tra i nuclei positivi, generando una forza attrattiva che bilancia la repulsione nucleo–nucleo. L’equilibrio tra queste forze definisce una distanza caratteristica, la lunghezza di legame, alla quale l’energia del sistema è minima (Figura 01.08-08).
Non tutti gli atomi sono limitati a un solo legame covalente. Gli elementi prevalenti nella chimica biologica mostrano tendenze regolari: l’ossigeno, con sei elettroni nel guscio esterno, tende a condividere due elettroni e forma fino a due legami covalenti; l’azoto, con cinque elettroni di valenza, tipicamente forma tre legami; il carbonio, con quattro elettroni di valenza, stabilisce fino a quattro legami covalenti, condividendo quattro coppie elettroniche (Figura 01.08-05). In molte molecole, il fosforo e lo zolfo possono espandere la condivisione elettronica oltre l’ottetto in specie stabili a rilevanza biologica, come i fosfati e alcuni derivati solforati.
Quando un atomo forma più legami, la disposizione spaziale risultante non è casuale ma segue l’orientazione degli orbitali coinvolti. Ne emergono angoli di legame caratteristici, oltre a lunghezze ed energie di legame specifiche (Figura 01.08-09). Per il carbonio tetracoordinato, i legami puntano verso i vertici di un tetraedro regolare, conferendo alle molecole organiche una geometria tridimensionale definita. Questa organizzazione spaziale, combinata con la possibilità di formare legami singoli, doppi o tripli, genera una ricchissima varietà di architetture molecolari.
I legami covalenti non sono tutti equivalenti: la loro polarità dipende dalla differenza di elettronegatività tra gli atomi. Quando tale differenza è significativa, la coppia elettronica è più vicina all’atomo più elettronegativo, producendo un legame covalente polare; se la distribuzione è simmetrica, il legame è apolare. La polarità influenza profondamente le interazioni tra molecole, la solubilità e l’organizzazione delle macromolecole biologiche.
In alternativa alla condivisione, il completamento del guscio di valenza può avvenire tramite trasferimento di elettroni, con formazione di ioni di carica opposta che si attraggono elettrostaticamente: è il caso dei legami ionici (Figura 01.08-06). In ambiente cellulare acquoso, questi legami sono spesso schermati dalle molecole di solvente, mentre i legami covalenti, molto direzionali e con energie di legame elevate, definiscono l’ossatura stabile delle biomolecole.
La combinazione di numero di legami formabili, loro lunghezza ed energia, e l’orientazione imposta dagli orbitali, costituisce il principio organizzativo della chimica del carbonio. Ne derivano proprietà come la rigidità dei doppi legami, la libera rotazione attorno ai legami singoli e la possibilità di ibridazione degli orbitali (ad esempio configurazioni tetraedriche, planari trigonali o lineari), che spiegano la varietà strutturale delle molecole organiche e la loro capacità di svolgere funzioni complesse in ambito biologico (Figura 01.08-09).
Image Gallery

Image Gallery
Image Gallery

Image Gallery
Image Gallery
Nel legame covalente più comune, una singola coppia di elettroni è condivisa tra due atomi: si parla di legame singolo. In numerosi casi, tuttavia, gli atomi mettono in comune due o tre coppie elettroniche, generando rispettivamente legami doppi e tripli. L’aumento del numero di coppie condivise, cioè dell’ordine di legame, si accompagna a una riduzione della lunghezza di legame e a un incremento della sua energia di dissociazione; in termini generali vale la tendenza r_single > r_double > r_triple ed E_single < E_double < E_triple. Ciò si riflette sulla rigidità strutturale delle molecole e sulla loro reattività.
I legami multipli si compongono di un legame σ (sigma), che congiunge gli atomi lungo l’asse internucleare, e di uno o due legami π (pi greco), formati dalla sovrapposizione laterale di orbitali p. Il legame σ consente la rotazione libera attorno all’asse solo quando è presente da solo, come nei legami singoli; la presenza di un legame π, tipica del legame doppio, impone invece una barriera alla rotazione e congela l’orientazione relativa dei sostituenti (Figura 01.08-10). Questa restrizione conformazionale può generare isomeria geometrica (cis/trans o E/Z) e ha effetti decisivi sulla forma e sulla funzione di molte macromolecole.
La delocalizzazione degli elettroni in sistemi coniugati conferisce caratteristiche intermedie tra legame singolo e doppio ai legami coinvolti. Un caso paradigmatico è il benzene: gli elettroni π sono distribuiti su tutto l’anello a sei atomi di carbonio, dando luogo a una struttura aromaticamente stabilizzata in cui tutti i legami C–C hanno lunghezza ed energia molto simili, pari a un ordine di legame efficace vicino a 1,5. Tale delocalizzazione, oltre a spiegare l’elevata stabilità, condiziona la reattività e la geometria planare dell’anello:
- legame singolo: un’unica coppia condivisa, rotazione generalmente libera, lunghezza maggiore e minore energia di legame;
- legame doppio: due coppie condivise (σ + π), distanza più breve, maggiore energia e impedimento alla rotazione intorno all’asse del legame;
- legame triplo: tre coppie condivise (σ + 2π), il più corto e il più energetico della serie, con spiccata linearità e rigidità.
Image Gallery

Quando due elementi diversi sono uniti da un legame covalente, gli elettroni condivisi non sono necessariamente ripartiti in modo simmetrico. Questa asimmetria, dovuta alla differente tendenza degli atomi ad attrarre densità elettronica (elettronegatività, classicamente quantificata sulla scala di Pauling), conduce a legami covalenti polari. Ne risulta una separazione di carica parziale: l’atomo più elettronegativo acquisisce una parziale carica negativa (δ−), mentre il partner diventa parzialmente positivo (δ+). L’entità della polarità può essere descritta dal momento di dipolo \( \mu \), definito come \( \mu = q \cdot r \), dove q è la carica separata e r la distanza tra i centri di carica.
Tra gli elementi ricorrenti nei sistemi biologici, O e N sono relativamente più elettronegativi (circa 3,4 e 3,0 sulla scala di Pauling), mentre H lo è meno, e C ha un valore intermedio (circa 2,6). Ne discende che i legami O–H e N–H presentano marcata polarità, con H parzialmente positivo, come illustrato in (Figura 01.08-11). Il legame C–H, per via delle elettronegatività più simili di C e H (2,6 e 2,1), è invece debolmente polare o spesso considerato quasi apolare in molti contesti molecolari. Legami come C=O o C≡N sono fortemente polari e contribuiscono significativamente al momento di dipolo complessivo delle molecole. La polarità dei legami influenza proprietà fisico-chimiche cruciali: solubilità in acqua, suscettibilità a interazioni non covalenti (per esempio, legami a idrogeno), spettroscopia e reattività. L’acqua stessa possiede un elevato momento di dipolo (circa 1,85 D), che spiega la sua efficacia come solvente di specie polari e ioniche:
- elettronegatività: misura della capacità di attrarre elettroni di legame, con andamento crescente da sinistra a destra e dall’alto verso il basso nella tavola periodica;
- legami polari: generano dipoli locali e cariche parziali, favorendo interazioni con solventi polari e legami a idrogeno;
- legami non polari o debolmente polari: distribuzione elettronica più uniforme, minore affinità per solventi polari, maggiore affinità per fasi apolari.
Image Gallery

Ogni legame covalente è caratterizzato da una lunghezza e da un’energia di dissociazione specifiche (Figura 01.08-10). La forza di legame si quantifica come l’energia necessaria a rompere una mole di legami identici, espressa in kilocalorie per mole (kcal/mol) o kilojoule per mole (kJ/mol), con la relazione di conversione \( 1\,\text{kcal/mol} \approx 4,184\,\text{kJ/mol} \). Valori tipici includono, a titolo esemplificativo, C–C singolo ~ 83 kcal/mol (≈ 347 kJ/mol) e C=C ~ 146 kcal/mol (≈ 611 kJ/mol).
Confrontate con l’energia termica disponibile a temperatura ambiente, \( RT \approx 2,5\,\text{kJ/mol} \) (≈ 0,6 kcal/mol) a 298 K, le energie dei legami covalenti risultano superiori di ordini di grandezza. Per questo motivo, le vibrazioni e le collisioni molecolari che avvengono nelle cellule non sono in grado, da sole, di scindere in modo indiscriminato i legami covalenti. La loro rottura avviene invece nell’ambito di reazioni chimiche specifiche, in cui catalizzatori biologici altamente selettivi, gli enzimi, abbassano le barriere di attivazione senza alterare l’energia intrinseca dei legami coinvolti:
- energia di legame: misura della stabilità del legame, proporzionale alla profondità della buca di potenziale associata;
- unità di misura: kcal/mol e kJ/mol, con \( 1\,\text{kcal} \simeq 4,184\,\text{kJ} \);
- confronto con l’energia termica: \( RT \) a 298 K è circa 0,6 kcal/mol, molto inferiore alle energie dei legami covalenti, a garanzia della loro persistenza nelle condizioni fisiologiche.
Image Gallery
Quando la differenza di elettronegatività tra due elementi è molto marcata, la condivisione degli elettroni viene sostituita dal loro trasferimento completo verso l’atomo più elettronegativo. Si formano così ioni di carica opposta che si attraggono per via elettrostatica costituendo un legame ionico. Un esempio classico è l’interazione tra sodio e cloro: l’atomo di sodio (Na) perde il suo elettrone di valenza diventando Na⁺, mentre il cloro (Cl) acquista un elettrone trasformandosi in Cl⁻; le specie risultanti si attraggono e, in fase cristallina, danno luogo alla struttura ordinata del cloruro di sodio, il comune sale da cucina (Figura 01.08-12). In un cristallo millimetrico sono presenti in modo regolare circa \( 2 \times 10^{19} \) cationi e altrettanti anioni (Figura 01.08-12).
La stabilità dell’interazione coulombiana tra due cariche q1 e q2 separate da una distanza r in un mezzo con costante dielettrica relativa \( \varepsilon_r \) può essere espressa da: \[ E = \frac{1}{4\pi \varepsilon_0 \varepsilon_r} \cdot \frac{q_1 q_2}{r}. \] In acqua, dove \( \varepsilon_r \) è elevata (circa 78 a 25 °C), l’energia di interazione è significativamente schermata; di conseguenza, molti sali (incluso NaCl) si dissolvono, liberando ioni isolati circondati da gusci di solvatazione formati da molecole d’acqua orientate dal campo elettrico ionico. In soluzione acquosa, le interazioni ioniche risultano tipicamente 10–100 volte più deboli dei legami covalenti, pur rimanendo essenziali per l’organizzazione e la funzione biologica.
Gli ioni positivi sono detti cationi, quelli negativi anioni. Piccoli ioni inorganici come Na⁺, K⁺, Cl⁻ e Ca²⁺ sono determinanti per vari processi: stabiliscono potenziali di membrana, modulano l’attività di numerosi enzimi, partecipano alla trasduzione del segnale e contribuiscono agli equilibri osmotici e acido–base:
- formazione: trasferimento elettronico che produce specie cariche opposte e attrazione elettrostatica risultante;
- fase solida: reticoli cristallini con bilanciamento rigoroso delle cariche e alta regolarità strutturale;
- in acqua: forte solvatazione e dissociazione in ioni liberi grazie all’elevata costante dielettrica del solvente;
- rilevanza biologica: contributo a potenziale di membrana, segnalazione (es. Ca²⁺), catalisi e omeostasi ionica.
Image Gallery
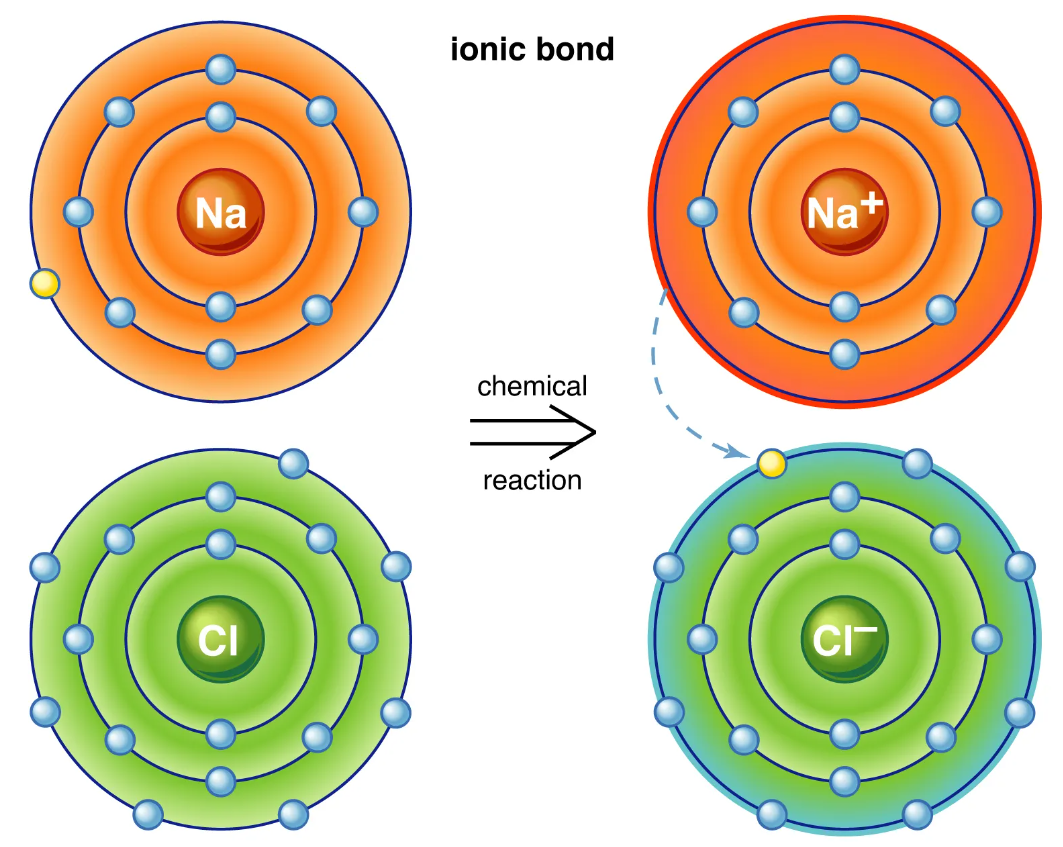
Image Gallery
L’acqua rappresenta in media circa il 70% della massa cellulare e funge da matrice per la grande maggioranza delle reazioni intracellulari. La chimica dei sistemi viventi è quindi modellata in modo decisivo dalle peculiari proprietà di H₂O. In ciascuna molecola di acqua, due atomi di idrogeno sono connessi all’ossigeno tramite legami covalenti O–H altamente polari: l’elevata elettronegatività dell’ossigeno attira la densità elettronica, lasciando sugli idrogeni una parziale carica positiva e sull’ossigeno una parziale carica negativa. La separazione di carica conferisce alla molecola un dipolo netto.
Quando una regione parzialmente positiva di una molecola d’acqua (un atomo di H) si avvicina alla regione parzialmente negativa di un’altra (l’atomo di O), si instaura un’attrazione elettrostatica direzionale, il legame a idrogeno (Figura 01.08-13). Si tratta di un’interazione nettamente più debole di un legame covalente e, in acqua liquida, la sua durata è dell’ordine di pochi picosecondi, poiché i moti termici ne favoriscono la rottura e il riformarsi continuo. Nonostante la debolezza del singolo legame, l’effetto collettivo di una rete estesa di legami a idrogeno è notevole: ogni molecola può agire contemporaneamente da donatrice e accettrice, generando un reticolo dinamico.
La rete di legami a idrogeno spiega perché l’acqua, a temperatura ambiente, è liquida e non gassosa, e perché mostra valori elevati di punto di ebollizione e tensione superficiale. A queste si affiancano un significativo calore specifico e un’elevata entalpia di vaporizzazione, aspetti che stabilizzano la temperatura negli ambienti biologici. Legami a idrogeno non si limitano però ai solventi: essi emergono ovunque un idrogeno parzialmente positivo, vincolato da un legame covalente polare, si trovi in prossimità di un accettore di coppie elettroniche con carica parziale negativa, come ossigeno o azoto appartenenti alla stessa o a un’altra molecola (Figura 01.08-13). Nelle macromolecole, legami a idrogeno intramolecolari contribuiscono alla stabilità conformazionale (per esempio, nelle strutture secondarie proteiche e nell’appaiamento di basi negli acidi nucleici), mentre quelli intermolecolari promuovono il riconoscimento specifico fra partner biologici.
Composti portatori di legami polari e capaci di formare legami a idrogeno sono detti idrofili: interagiscono favorevolmente con H₂O e risultano miscibili o solubili. In questa categoria ricadono zuccheri, DNA, RNA e molte proteine. Al contrario, molecole prive di cariche o di dipoli significativi sono idrofobe e tendono a escludere l’acqua; specie anfipatiche, che combinano porzioni polari e non polari, mostrano un comportamento intermedio, fondamentale per l’auto-assemblaggio di membrane e micelle.
Image Gallery

Image Gallery
Numerosi eventi biologici dipendono da contatti transitori ma selettivi tra molecole. Tali associazioni sono governate da interazioni non covalenti, la cui efficacia deriva dalla somma cooperativa di contributi individualmente modesti. Oltre ai legami a idrogeno, rivestono un ruolo centrale le attrazioni elettrostatiche, le forze di van der Waals e l’effetto idrofobico.
Le attrazioni elettrostatiche comprendono le interazioni tra cariche opposte, come quelle che uniscono Na⁺ e Cl⁻ in un cristallo ionico (Figura 01.08-12). Possono manifestarsi anche tra regioni parzialmente cariche di molecole con legami covalenti polari (Figura 01.08-11). L’intensità dell’attrazione tra due cariche puntiformi è descritta da \( F = \frac{1}{4\pi \varepsilon_0 \varepsilon_r} \frac{q_1 q_2}{r^2} \): in acqua, l’elevato coefficiente dielettrico \( \varepsilon_r \) attenua marcatamente le forze coulombiane, riducendo la portata delle interazioni a distanza. Ciononostante, su superfici biomolecolari estese, la disposizione complementare di molteplici cariche e dipoli può dare luogo a riconoscimenti molto specifici e stabili (Figura 01.08-14).
Le attrazioni di van der Waals emergono quando atomi neutri si avvicinano a distanze inferiori alla somma dei loro raggi di van der Waals. Fluttuazioni istantanee nella densità elettronica generano dipoli temporanei che si inducono reciprocamente, producendo un’attrazione a corto raggio. Queste interazioni, deboli e aspecifiche, sono ubiquitarie e agiscono anche su molecole prive di cariche formali o di forte polarità. Le lunghezze e i valori energetici relativi dei legami non covalenti appena discussi sono riportati in confronto ai legami covalenti nella (Tabella 01.08-01).
Il quarto contributo, spesso determinante nei sistemi acquosi, è l’effetto idrofobico. Non si tratta di un vero legame, ma della tendenza delle superfici non polari a segregarsi dall’acqua per minimizzare la perturbazione della rete di legami a idrogeno del solvente. In altre parole, l’aggregazione di gruppi apolari riduce l’area di contatto con H₂O, rendendo più favorevole l’assetto entropico complessivo. Questo principio è cruciale per l’auto-organizzazione delle membrane cellulari: lipidi con lunghe catene idrocarburiche, i cui legami C–H sono pressoché apolari, non stabiliscono legami a idrogeno con l’acqua e, perciò, si dispongono in doppi strati che delimitano un compartimento acquoso interno separandolo dall’esterno:
- legami a idrogeno: direzionali, dipolo–dipolo, tipicamente tra donatori X–H e accettori Y con geometrie prossime alla linearità;
- attrazioni elettrostatiche: tra cariche formali o parziali, schermate dal solvente in funzione di \( \varepsilon_r \) e della forza ionica;
- forze di van der Waals: interazioni di dispersione e induzione a corto raggio, massime a distanze prossime alla somma dei raggi di van der Waals;
- effetto idrofobico: aggregazione di superfici apolari in acqua, guidata dalla minimizzazione della perturbazione della rete H₂O e dal guadagno entropico del solvente.
Image Gallery
Image Gallery
Image Gallery

Lunghezza e stabilità dei principali legami chimici
| Tipo di legame | Lunghezza (nm) | Forza (kJ/mol) nel vuoto | Forza (kJ/mol) in acqua |
|---|---|---|---|
| Covalente | 0,10 | 377 (90 kcal/mol) | 377 (90 kcal/mol) |
| Non covalente: legame ionico | 0,25 | 355 (85 kcal/mol) | 126 (3 kcal/mol) |
| Non covalente: legame idrogeno | 0,17 | 16,7 (4 kcal/mol) | 4,2 (1 kcal/mol) |
| Non covalente: interazioni di van der Waals | 0,35 | 0,4 (0,1 kcal/mol) | 0,4 (0,1 kcal/mol) |
| Tabella sulla lunghezza e stabilità dei principali legami chimici. | |||
Quando una molecola dotata di un legame X–H fortemente polare entra in soluzione, l’idrogeno può risultare fortemente deprivato di densità elettronica e comportarsi come un protone quasi nudo, H⁺. Le molecole d’acqua circostanti, con atomi di ossigeno parzialmente negativi, solvatano il protone formando ioni idronio, H₃O⁺ (Figura 01.08-11) e (Figura 01.08-15). L’associazione e la dissociazione del protone sono processi estremamente rapidi e reversibili, dando luogo a un equilibrio dinamico nel quale i protoni “saltano” fra specie adiacenti.
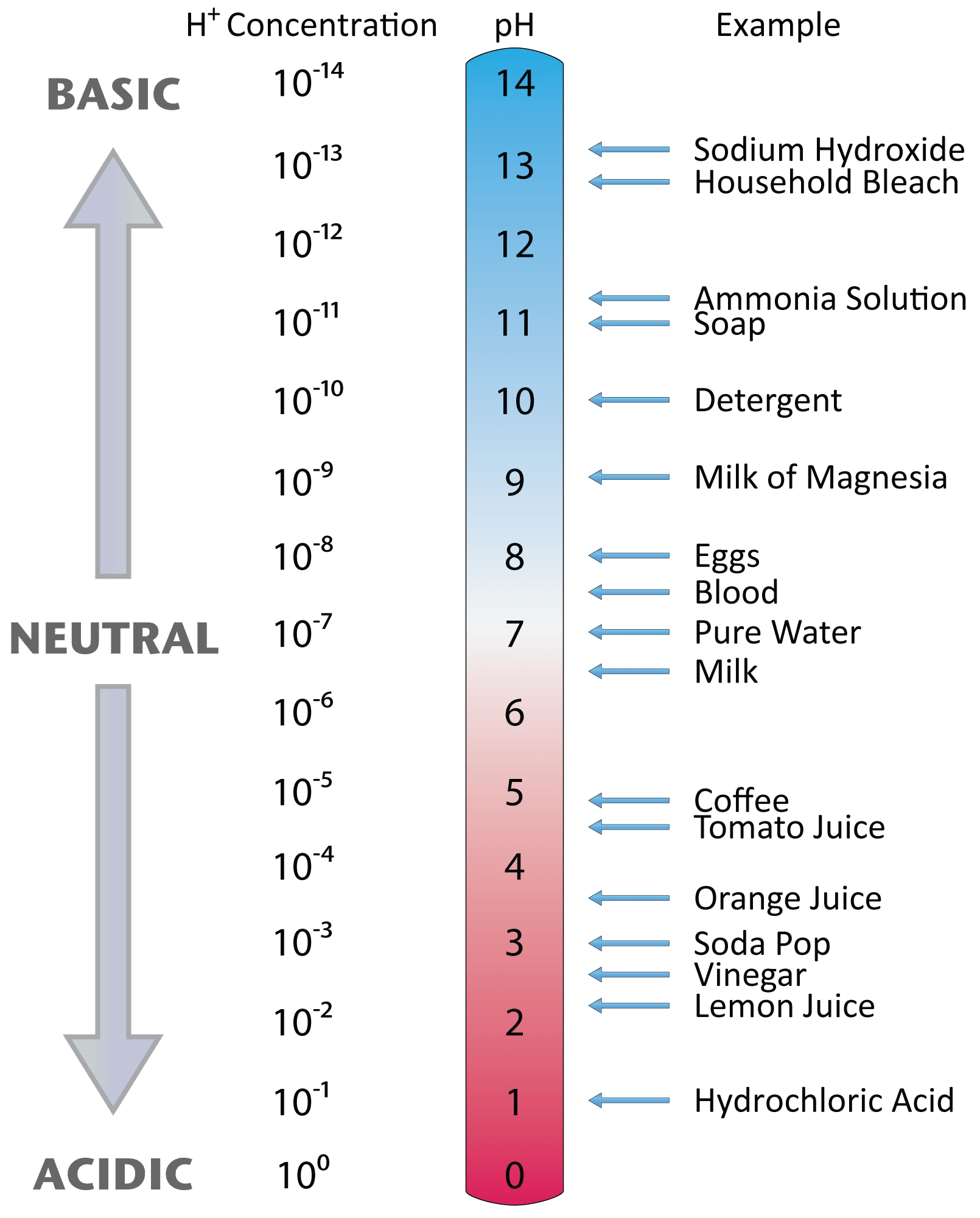
Nell’acqua pura è presente una lieve autoionizzazione che genera concentrazioni identiche di H₃O⁺ e OH⁻ pari a 10⁻⁷ M (Figura 01.08-15). Il prodotto ionico dell’acqua è \( K_w = 1,0 \times 10^{-14} \) a 25 °C, da cui derivano le relazioni \( \mathrm{pH} = -\log[ \mathrm{H}^+ ] \), \( \mathrm{pOH} = -\log[ \mathrm{OH}^- ] \) e \( \mathrm{pH} + \mathrm{pOH} = 14,0 \). Per convenzione, si parla di soluzione acida se pH < 7,0, basica (o alcalina) se pH > 7,0, e neutra a pH 7,0.
Un acido, in senso di Brønsted–Lowry, è un donatore di protoni; una base è un accettore di protoni. Gli acidi forti cedono H⁺ quasi completamente in acqua (per esempio, l’acido nitrico), mentre gli acidi deboli presentano dissociazioni parziali, quantificate da \( K_a \) e dal corrispondente \( \mathrm{p}K_a = -\log K_a \). Molti gruppi carbossilici (–COOH) in biomolecole si comportano da acidi deboli. Per le basi, specie forti come KOH liberano rapidamente OH⁻, mentre basi deboli – tra cui ammine biologicamente rilevanti – accettano un protone dall’acqua secondo \( \mathrm{R{-}NH_2} + \mathrm{H_2O} \rightleftharpoons \mathrm{R{-}NH_3^+} + \mathrm{OH^-} \). L’aumento di [OH⁻] riduce [H⁺] e viceversa, mantenendo \( [\mathrm{H}^+]\,[\mathrm{OH}^-] = K_w \) (Figura 01.08-16).
La sensibilità delle specie debolmente acide o basiche alle variazioni di pH è funzionale alla regolazione biologica: piccole modifiche della protonazione possono cambiare carica, solubilità e reattività di proteine e metaboliti. Nella cellula, il pH citosolico si mantiene vicino alla neutralità (circa 7,0–7,2) grazie a sistemi tampone, miscele di un acido debole e della base coniugata che resistono alle variazioni di pH. La relazione di Henderson–Hasselbalch,
\( \mathrm{pH} = \mathrm{p}K_a + \log \left( \frac{[\text{base coniugata}]}{[\text{acido}]} \right) \),
descrive la dipendenza del pH dal rapporto fra le due componenti. Per esempio, un tampone fosfato H₂PO₄⁻/HPO₄²⁻ con \( \mathrm{p}K_a \approx 6,86 \) mantiene un pH prossimo alla neutralità: se il rapporto [HPO₄²⁻]/[H₂PO₄⁻] è 2,0, allora \( \mathrm{pH} \approx 6,86 + \log(2,0) = 7,16 \). Sistemi tampone intracellari (fosfati, istidina in proteine) e, a livello sistemico, il bicarbonato sono esempi chiave del controllo del pH. Il bilanciamento dinamico tra rilascio e acquisizione di protoni da parte di acidi e basi deboli stabilizza così l’ambiente chimico cellulare attorno a pH 7,0.
Image Gallery
Image Gallery

Image Gallery
Image Gallery