


Le mutazioni di uno o pochi nucleotidi possono influenzare la struttura e la funzione di una proteina
TOPICS
Definizione

I recettori accoppiati a enzimi, analogamente ai recettori accoppiati a proteine G, sono proteine transmembrana che presentano il sito di legame per il ligando sulla superficie esterna della membrana plasmatica. A differenza dei recettori accoppiati a proteine G, tuttavia, il dominio citoplasmatico di questi recettori possiede direttamente un’attività catalitica oppure associa, in modo stabile e regolato, un enzima citosolico con funzione definita. La loro importanza emerse dallo studio delle risposte a proteine segnale extracellulari, in gran parte fattori di crescita e citochine, che modulano crescita, proliferazione, differenziamento e sopravvivenza cellulare nei tessuti animali (Tabella 05.25-01). Questi mediatori agiscono spesso in prossimità del sito di secrezione e a concentrazioni estremamente basse, tipicamente nell’intervallo picomolare-nanomolare, ossia circa da \(10^{-11}\) a \(10^{-9}\) M. Le risposte da loro evocate sono per lo più lente, richiedono ore, e coinvolgono cascate intracellulari che convergono su modificazioni dell’espressione genica. Nonostante il ruolo centrale nella regolazione trascrizionale, i recettori accoppiati a enzimi possono innescare anche rimodellamenti rapidi dell’architettura citoscheletrica, con effetti immediati su forma cellulare e motilità. In tali contesti, il segnale extracellulare non è sempre una proteina solubile; di frequente si tratta di proteine ancorate alla matrice o alla superficie di cellule adiacenti, che fungono da segnali di contatto durante l’adesione e la migrazione. La famiglia più ampia di recettori accoppiati a enzimi è costituita dai recettori tirosina chinasi (RTK, Receptor Tyrosine Kinase), nei quali il dominio citoplasmatico esercita un’attività chinasica specifica per residui di tirosina su substrati intracitoplasmatici selezionati. Nei paragrafi successivi verranno considerati i principi di attivazione degli RTK in risposta a segnali extracellulari, la propagazione del segnale lungo due direttrici principali che confluiscono su molteplici proteine effettrici, e i casi in cui recettori accoppiati a enzimi deviano da queste vie per modulare più direttamente la trascrizione. Poiché anomalie della segnalazione mediata da RTK e recettori affini promuovono crescita, sopravvivenza, differenziamento anomalo e invasività, alterazioni in tali circuiti sono frequentemente implicate nell’oncogenesi. Accanto agli RTK, rientrano tra i recettori accoppiati a enzimi anche: i recettori serina/treonina chinasi (ad es. recettori per TGF-β), i recettori guanilato ciclasi di membrana, i recettori tirosina fosfatasi e i recettori privi di attività catalitica intrinseca che reclutano chinasi citoplasmatiche non recettoriali (p.es. le chinasi della famiglia JAK nelle vie JAK-STAT). Tale varietà strutturale consente un’ampia gamma di logiche di segnalazione, dalla fosforilazione proteica a modifiche dei livelli di secondi messaggeri.
Molecole segnale e loro effetti biologici
| Molecola segnale | Origine | Natura chimica | Effetti principali |
|---|---|---|---|
| Adrenalina | Ghiandole surrenali | Derivato della tirosina | Aumenta pressione sanguigna, frequenza cardiaca e metabolismo |
| Cortisolo | Corticale surrenale | Steroide (colesterolo) | Regola metabolismo di carboidrati e proteine, risposta allo stress |
| Estradiolo | Ovaie | Steroide (colesterolo) | Stimola sviluppo dei caratteri sessuali femminili |
| Insulina | Cellule β pancreatiche | Proteina | Promuove ingresso di glucosio e sintesi di glicogeno e lipidi |
| Testosterone | Testicoli | Steroide (colesterolo) | Sviluppo caratteri sessuali maschili |
| Tiroxina | Tiroide | Derivato della tirosina | Regola metabolismo e differenziamento cellulare |
| Fattore di crescita dell’epidermide (EGF) | Varie cellule | Proteina | Stimola crescita e proliferazione cellulare |
| Fattore di crescita piastrinico (PDGF) | Piastrine e cellule varie | Proteina | Promuove divisione cellulare e cicatrizzazione |
| Fattore di crescita nervoso (NGF) | Tessuti nervosi | Proteina | Favorisce sopravvivenza e crescita di neuroni |
| Istamina | Mastociti | Derivato dell’istidina | Vasodilatazione e aumento permeabilità capillare |
| Ossido nitrico (NO) | Neuroni, cellule endoteliali | Gas | Rilassa muscolatura liscia e regola flusso sanguigno |
| Acetilcolina | Terminali nervosi | Derivato della colina | Neurotrasmettitore eccitatorio in sinapsi e giunzioni neuromuscolari |
| GABA | Neuroni | Derivato dell’acido glutammico | Neurotrasmettitore inibitorio |
| Delta (segnale contatto-dipendente) | Cellule in sviluppo | Proteina transmembrana | Controlla differenziamento cellulare in fase embrionale |
| Tabella che descrive il ruolo delle molecole segnale nella regolazione di comunicazioni complesse tra cellule e tessuti, includendo ormoni, mediatori locali, neurotrasmettitori e segnali contatto-dipendenti, responsabili della modulazione di funzioni vitali come metabolismo, crescita, differenziamento e attività neuronale, assicurando la coordinazione dei processi fisiologici dell’organismo. | |||
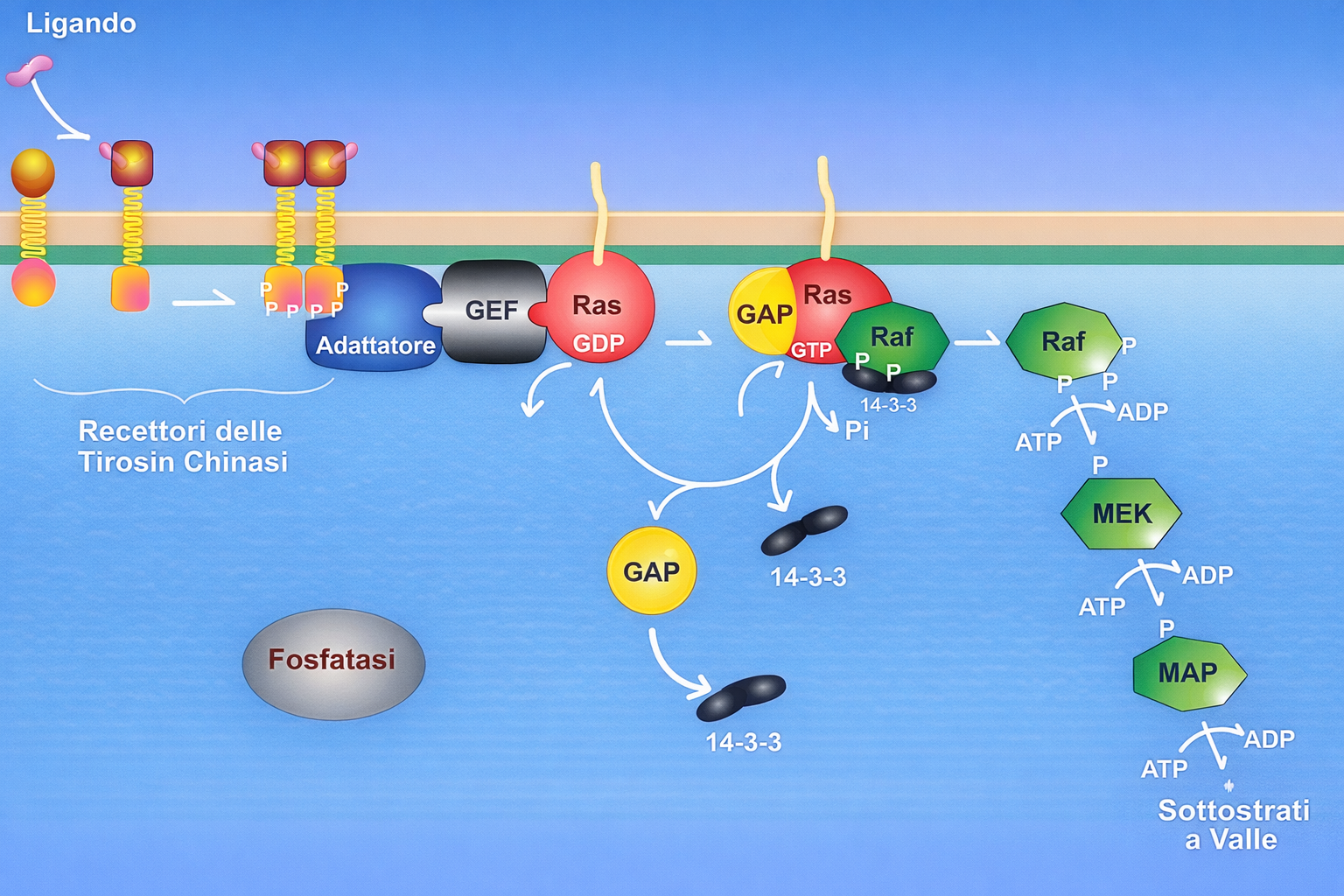
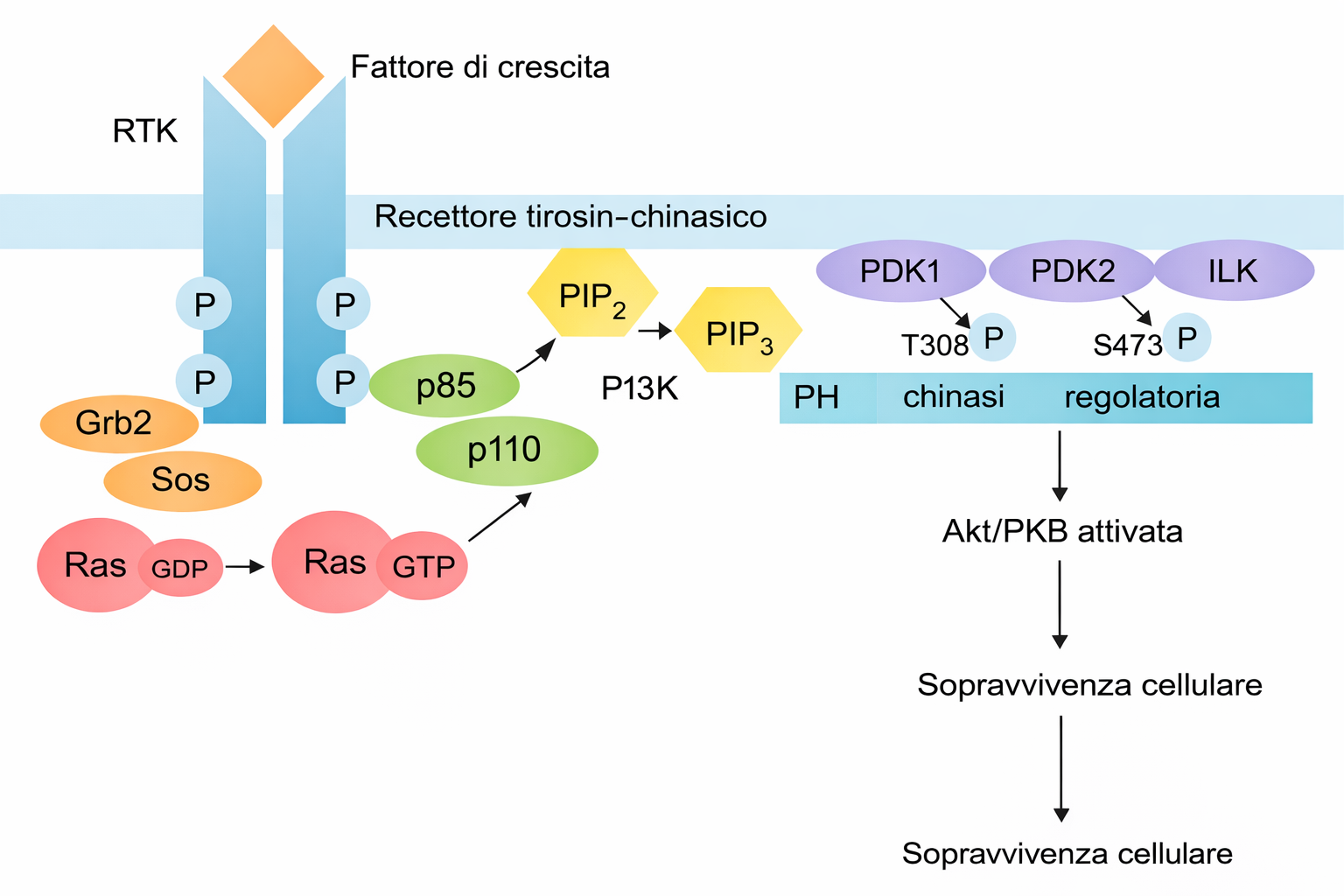
Affinché un recettore accoppiato a enzimi svolga la funzione di trasduttore, l’interazione del ligando con il dominio extracellulare deve tradursi nell’attivazione del modulo enzimatico intracellulare (o dell’enzima a esso associato). A differenza dei GPCR, che attraversano la membrana sette volte, i recettori accoppiati a enzimi possiedono in genere un singolo segmento transmembrana α-elicoidale. Poiché un’unica elica non è efficiente nel trasmettere grandi riarrangiamenti conformazionali attraverso il doppio strato, l’attivazione si realizza prevalentemente tramite cambiamenti nello stato di oligomerizzazione. Nella maggioranza degli RTK, il legame del ligando promuove l’avvicinamento di due monomeri e la formazione di un dimero, che può essere stabilizzato dal ligando stesso (dimerizzazione mediata dal ligando) o da domini extracellulari predisposti all’interazione. L’accostamento delle code citoplasmatiche consente l’autofosforilazione in trans del dominio chinasico, innescando l’attività catalitica e aprendo i siti di docking su residui di tirosina specifici. La fosforilazione delle tirosine nel dominio citosolico del recettore crea siti di legame ad alta affinità per un repertorio di proteine di segnalazione che possiedono domini di riconoscimento di fosfo-tirosina, quali SH2 e PTB. Sulle code degli RTK si organizza quindi un complesso di trasduzione costituito da numerosi componenti, spesso dell’ordine di 10–20 specie proteiche (Figura 05.25-01). Alcune vengono esse stesse fosforilate e attivate dal recettore, propagando il segnale; altre funzionano da adattatori o piattaforme di assemblaggio, collegando il recettore ad ulteriori nodi della rete. Meccanismi analoghi consentono a proteine di segnalazione di riconoscere lipidi fosforilati prodotti nella membrana in risposta a stimoli, creando ulteriori siti di reclutamento e compartimentalizzazione della risposta. Una volta costituiti, questi complessi convertono il segnale in una risposta coordinata attivando simultaneamente più vie che raggiungono bersagli distinti all’interno della cellula. Ciò permette di integrare programmi funzionali complessi, come proliferazione o differenziamento, che richiedono il coordinamento di molteplici effettori. La durata e l’ampiezza della risposta dipendono da meccanismi di spegnimento che includono:
- defosforilazione dei residui di tirosina del recettore e dei componenti del segnale a opera di proteina tirosina fosfatasi;
- endocitosi mediata da clatrina o caveole degli RTK attivati, spesso preceduta da ubiquitinazione dipendente da E3 ligasi, con successivo riciclo o degradazione lisosomiale, processo condiviso anche da alcuni GPCR;
- feedback negativi intracitoplasmatici, ad esempio induzione di inibitori o fosfatasi specifiche che limitano la propagazione del segnale.
Sebbene diversi RTK reclutino insiemi distinti di effettori, alcuni moduli ricorrono con alta frequenza. Tra questi figurano:
- la fosfolipasi C-γ, che opera in modo strettamente analogo alla fosfolipasi C attivata dai GPCR nella via dei fosfoinositidi, generando diacilglicerolo e IP3 e promuovendo l’attivazione della proteina chinasi C e il rilascio di Ca2+ dai depositi intracellulari;
- le vie Ras–MAP chinasi e PI3K–Akt, frequentemente attivate a valle degli RTK e responsabili rispettivamente di programmi proliferativi/differenziativi e di segnali di crescita e sopravvivenza;
- adattatori citosolici quali Grb2, Shc e scaffold modulari, che collegano i siti fosfo-tirosinici del recettore alle GTPasi della famiglia Ras e ad altre chinasi a valle.
Oltre alle risposte trascrizionali, gli RTK e altri recettori accoppiati a enzimi possono indurre riconfigurazioni rapide del citoscheletro, modulando GTPasi della famiglia Rho, complessi di adesione e dinamiche di actina; in molti casi lo stimolo è fornito da proteine di membrana o di matrice non diffusibili che fungono da segnali spazialmente ristretti durante la migrazione cellulare (Figura 05.25-02). La corretta regolazione temporale e spaziale di queste vie è essenziale per l’omeostasi tissutale; mutazioni attivanti nei recettori o nei loro effettori, così come perdita di controlli negativi, determinano iperattività del segnale con esiti patologici, tra cui numerose neoplasie. La natura modulare delle piattaforme di segnalazione su RTK spiega, inoltre, l’efficacia clinica di inibitori mirati delle chinasi, di anticorpi contro i recettori e di agenti che interferiscono con l’endocitosi o la degradazione recettoriale (Figura 05.25-03).
Image Gallery
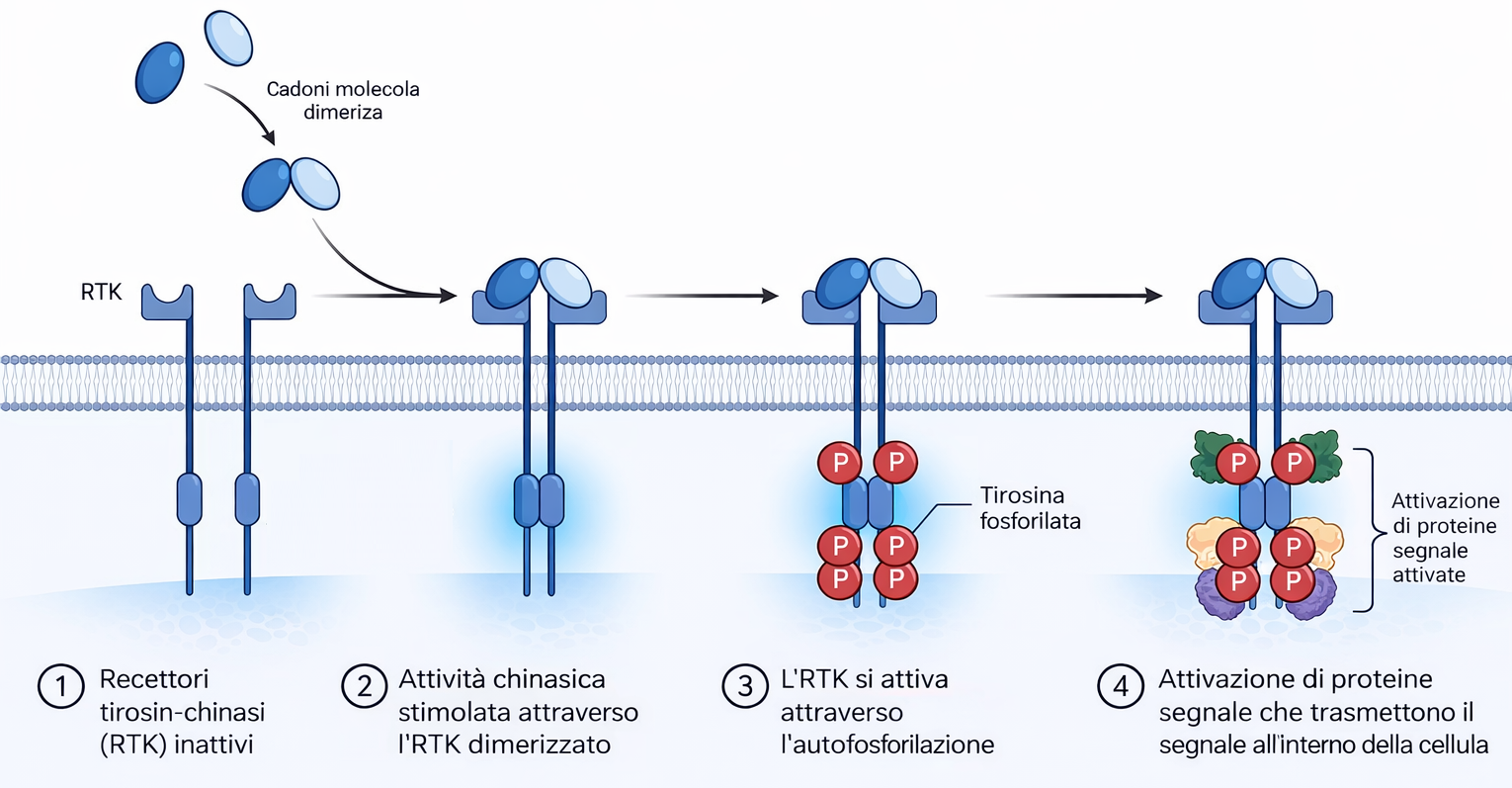
Image Gallery
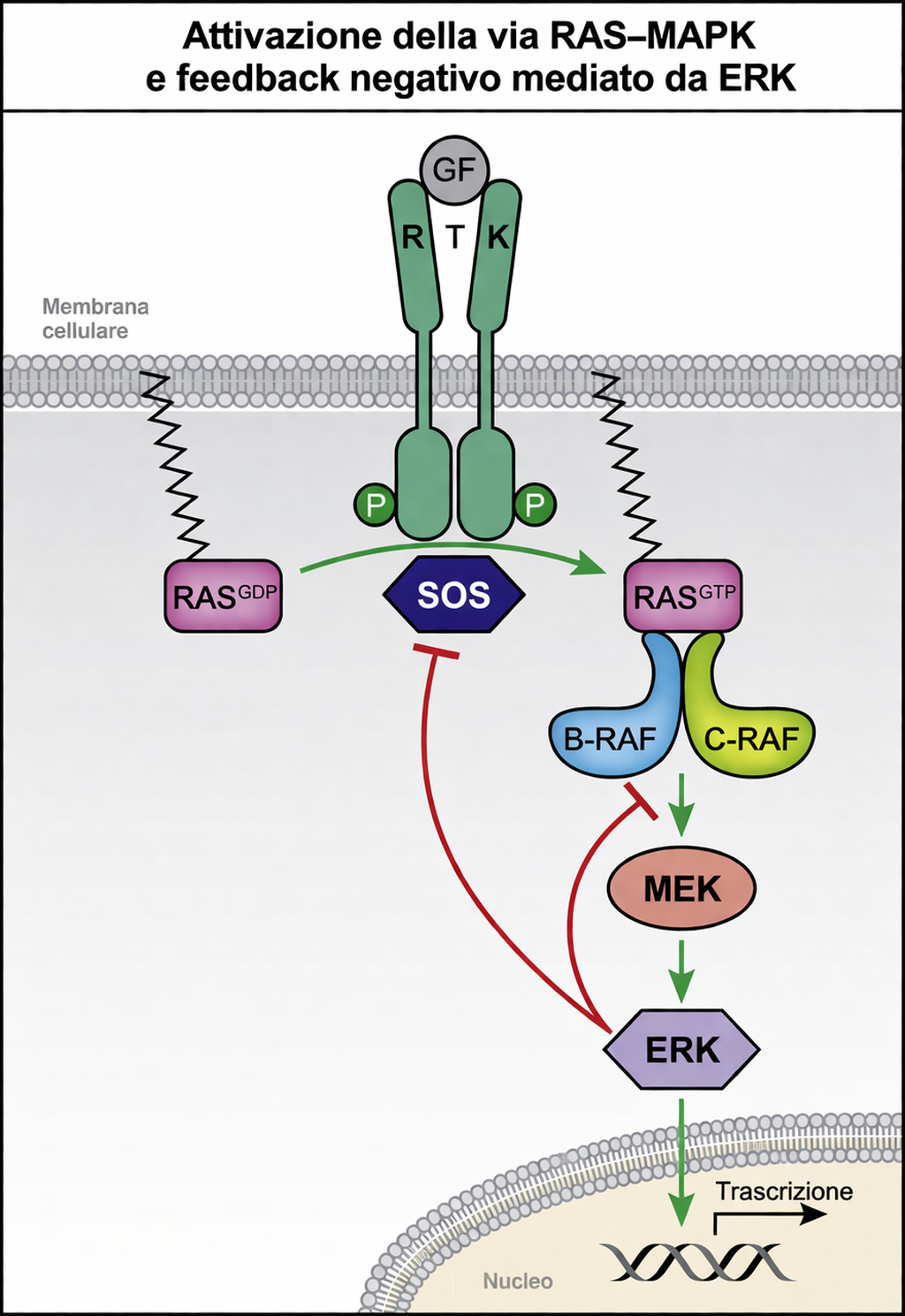
Image Gallery
Non tutti i recettori accoppiati a enzimi demandano il messaggio a lunghe cascate citoplasmatiche; alcuni instradano l’informazione verso il nucleo con una via quasi diretta. Un paradigma è Notch, un recettore essenziale negli animali, attivo sia in fase embrionale sia in età adulta, che in Drosophila governa, tra l’altro, la selezione delle cellule nervose per inibizione laterale (Figura 05.25-06). In questo sistema, il recettore stesso contribuisce alla regolazione trascrizionale. Il legame di ligandi di membrana come Delta, esposti sulla cellula adiacente, induce clivaggi proteolitici sequenziali del recettore: la porzione intracitoplasmatica viene rilasciata e trasloca nel nucleo, dove coopera con fattori di trascrizione per modulare un set definito di geni bersaglio sensibili a Notch). Meccanicisticamente, l’attivazione dipende da un taglio extracellulare mediato da metalloproteasi di tipo ADAM e da una scissione intramembrana catalizzata dal complesso γ-secretasi; il frammento nucleare (NICD) si associa a proteine della famiglia CSL, reclutando coattivatori e convertendo complessi repressivi in attivi. Questa “corsia preferenziale” consente di collegare contatti cellula‑cellula a programmi trascrizionali con notevole rapidità, minimizzando l’intermediazione citoplasmatica (Figura 05.25-04).
Image Gallery
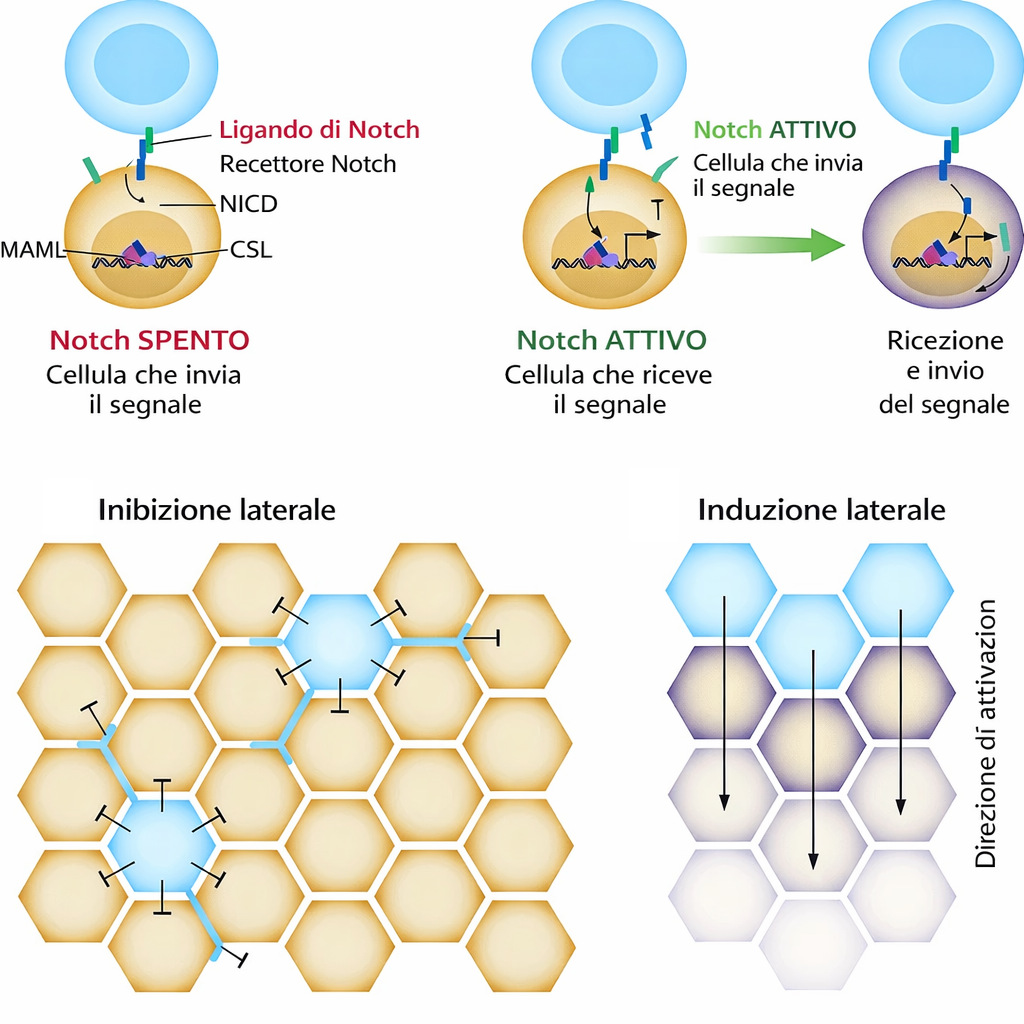
Image Gallery
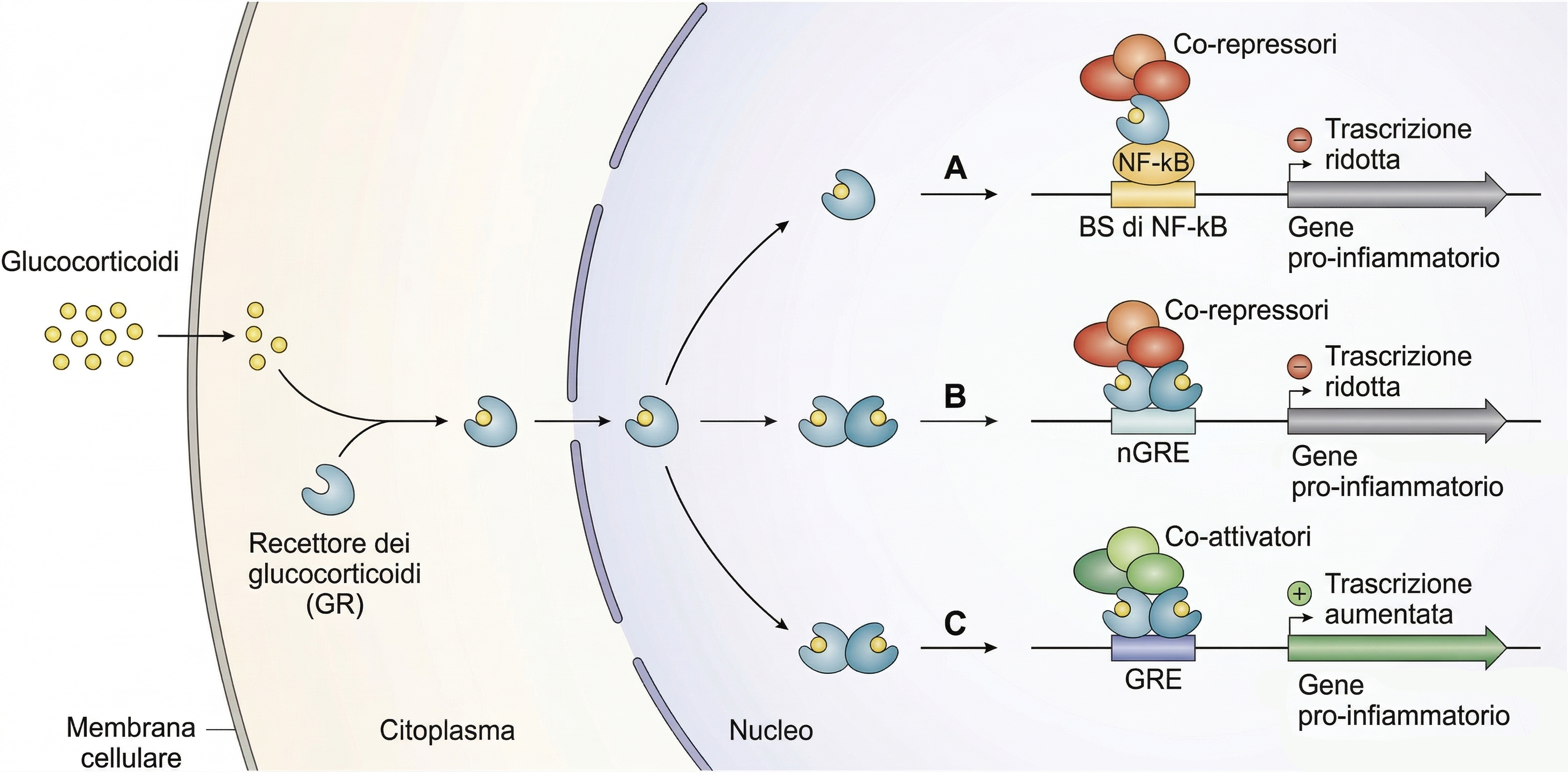
Una seconda via breve verso il nucleo è percorsa da molecole segnale idrofobe che utilizzano recettori intracellulari. Tra queste rientrano gli ormoni steroidei, come cortisolo, estradiolo e testosterone, e gli ormoni tiroidei, tra cui la tiroxina T4 (Figura 05.25-07). Queste molecole attraversano la membrana plasmatica delle cellule bersaglio e legano recettori situati nel citosol o direttamente nel nucleo. Tali proteine recettoriali, considerate complessivamente “recettori nucleari”, una volta attivate modulano l’espressione genica in maniera specifica (Figura 05.25-05). In condizioni basali i recettori nucleari sono inattivi, spesso associati a chaperoni o corepressori. Il legame dell’ormone induce un pronunciato rimodellamento conformazionale che abilita dimerizzazione, legame a elementi di risposta sul DNA e reclutamento di cofattori che attivano o reprimono la trascrizione (Figura 05.25-08). Specificità e diversità nascono da tre dimensioni complementari: selettività di legame ligando‑recettore, riconoscimento di distinti siti regolativi sul genoma e contesto cellulare, che determina il repertorio di cofattori e lo stato della cromatina. Così, il medesimo ormone può orchestrare insiemi genici differenti in cellule diverse, generando risposte fisiologiche non sovrapponibili. I recettori nucleari e i loro ligandi hanno un impatto sistemico nella fisiologia umana (Tabella 05.25-01). Alterazioni genetiche in questi assi di segnalazione comportano fenotipi marcati. Un esempio è la sindrome da insensibilità agli androgeni, dovuta a varianti inattivanti del recettore del testosterone: in individui con assetto cromosomico XY, la produzione di androgeni è presente ma le cellule non possono tradurre il segnale. Nella forma completa, i genitali esterni assumono morfologia femminile, con assenza di risposta puberale ai segnali androgenici pur in presenza di livelli circolanti di ormone; nella forma parziale, compaiono quadri intermedi, a testimonianza di come un singolo recettore governi programmi trascrizionali in molti tessuti diversi, dal sistema riproduttivo al sistema nervoso.
Image Gallery
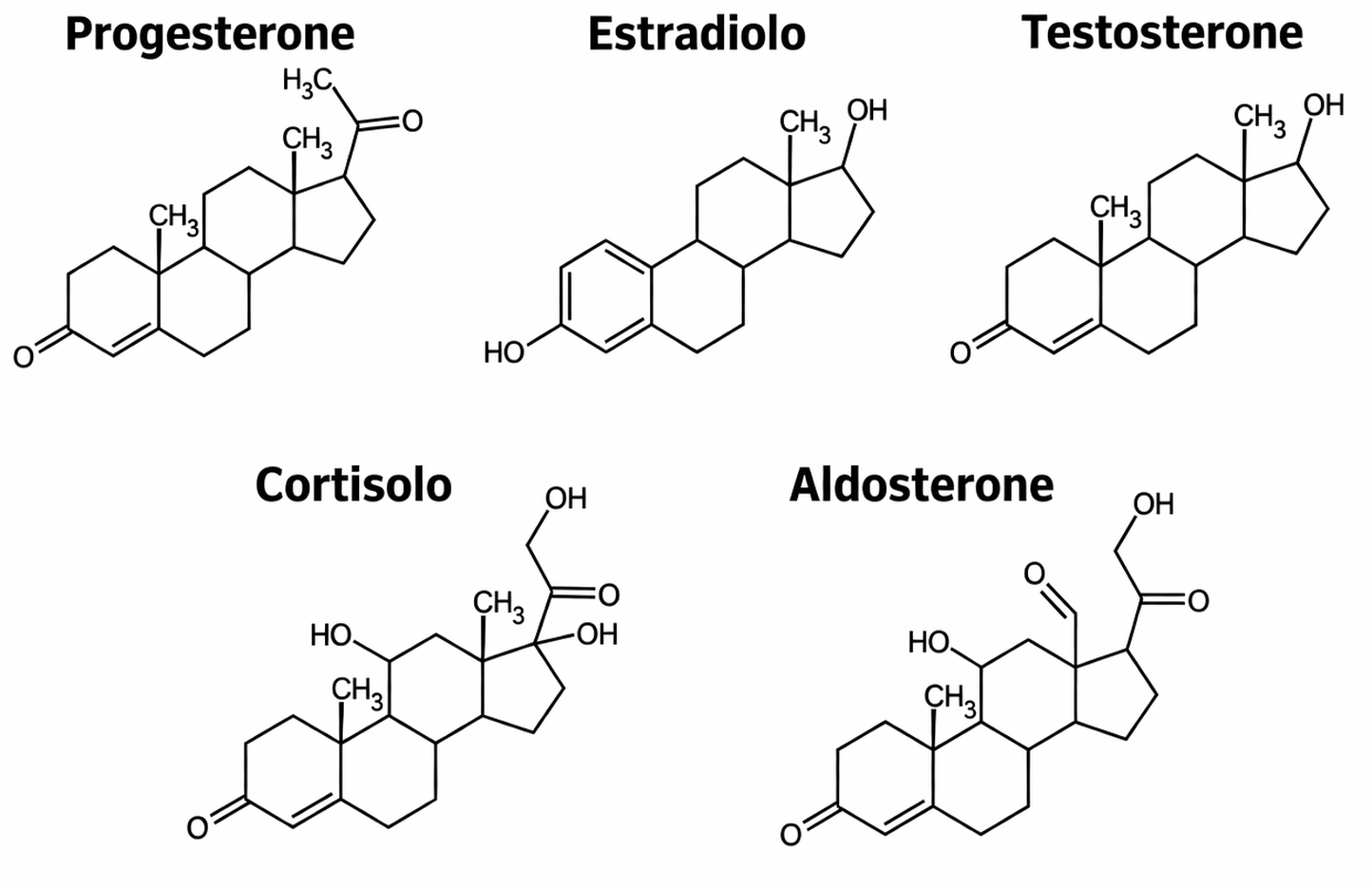
Image Gallery
Image Gallery
Molecole segnale e loro effetti biologici
| Molecola segnale | Origine | Natura chimica | Effetti principali |
|---|---|---|---|
| Adrenalina | Ghiandole surrenali | Derivato della tirosina | Aumenta pressione sanguigna, frequenza cardiaca e metabolismo |
| Cortisolo | Corticale surrenale | Steroide (colesterolo) | Regola metabolismo di carboidrati e proteine, risposta allo stress |
| Estradiolo | Ovaie | Steroide (colesterolo) | Stimola sviluppo dei caratteri sessuali femminili |
| Insulina | Cellule β pancreatiche | Proteina | Promuove ingresso di glucosio e sintesi di glicogeno e lipidi |
| Testosterone | Testicoli | Steroide (colesterolo) | Sviluppo caratteri sessuali maschili |
| Tiroxina | Tiroide | Derivato della tirosina | Regola metabolismo e differenziamento cellulare |
| Fattore di crescita dell’epidermide (EGF) | Varie cellule | Proteina | Stimola crescita e proliferazione cellulare |
| Fattore di crescita piastrinico (PDGF) | Piastrine e cellule varie | Proteina | Promuove divisione cellulare e cicatrizzazione |
| Fattore di crescita nervoso (NGF) | Tessuti nervosi | Proteina | Favorisce sopravvivenza e crescita di neuroni |
| Istamina | Mastociti | Derivato dell’istidina | Vasodilatazione e aumento permeabilità capillare |
| Ossido nitrico (NO) | Neuroni, cellule endoteliali | Gas | Rilassa muscolatura liscia e regola flusso sanguigno |
| Acetilcolina | Terminali nervosi | Derivato della colina | Neurotrasmettitore eccitatorio in sinapsi e giunzioni neuromuscolari |
| GABA | Neuroni | Derivato dell’acido glutammico | Neurotrasmettitore inibitorio |
| Delta (segnale contatto-dipendente) | Cellule in sviluppo | Proteina transmembrana | Controlla differenziamento cellulare in fase embrionale |
| Tabella che descrive il ruolo delle molecole segnale nella regolazione di comunicazioni complesse tra cellule e tessuti, includendo ormoni, mediatori locali, neurotrasmettitori e segnali contatto-dipendenti, responsabili della modulazione di funzioni vitali come metabolismo, crescita, differenziamento e attività neuronale, assicurando la coordinazione dei processi fisiologici dell’organismo. | |||
La linea evolutiva che conduce alle piante si è separata da quella degli animali più di un miliardo di anni fa, a partire da un progenitore eucariotico unicellulare. La transizione verso la pluricellularità è quindi avvenuta in modo indipendente, con l’emersione di soluzioni molecolari parzialmente convergenti e in parte divergenti. Una base genetica comune, ereditata da antenati unicellulari già capaci di comunicare, spiega alcune somiglianze; tuttavia, differenze sostanziali riflettono la storia evolutiva separata. Le piante impiegano ampiamente recettori di superficie di tipo chinasi serina/treonina, appartenenti a vaste famiglie di recettori simili a chinasi (RLK) in Arabidopsis thaliana, coinvolti nella crescita, nello sviluppo e nella risposta a patogeni. A differenza degli animali, nelle piante sono rari recettori GPCR canonici, non si osserva un ruolo centrale per AMP ciclico e mancano archetipi di RTK e recettori nucleari steroidei tipici del regno animale. Uno dei circuiti meglio caratterizzati è la risposta all’etilene, un ormone gassoso che regola eventi quali germinazione e maturazione dei frutti, sfruttato anche in post‑raccolta per modulare la maturazione del pomodoro. I recettori dell’etilene sono proteine di membrana che, pur funzionando come recettori accoppiati a chinasi, non sono imparentati con le classi descritte negli animali. Peculiare è la loro attività costitutiva in assenza di ligando: senza etilene il recettore attiva una chinasi a valle che sopprime la trascrizione dei geni di risposta nel nucleo; quando l’etilene si lega, recettore e chinasi vengono inattivati, liberando la trascrizione dei geni bersaglio (Figura 05.25-09). Questa logica di “de‑repressione” è ricorrente nei circuiti vegetali.
Image Gallery
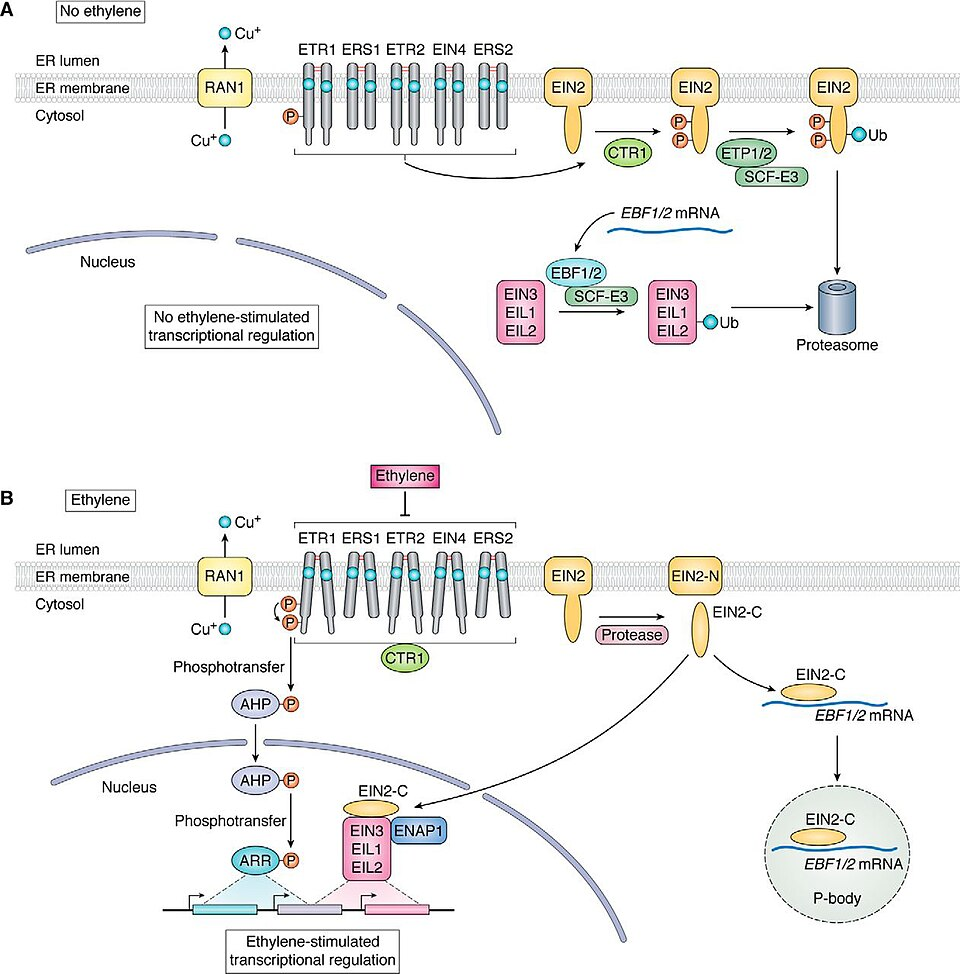
Sia nelle piante sia negli animali, le cellule integrano flussi simultanei di informazioni per decidere risultati complessi, come sopravvivenza o morte, proliferazione, differenziamento, rimodellamento morfologico, motilità e secrezione di mediatori. Questa integrazione nasce dalla fitta interconnessione tra vie di segnalazione, con estesa comunicazione incrociata. In tale rete, le proteine chinasi svolgono un ruolo nodale: oltre a fosforilare i componenti del proprio percorso, spesso modificano elementi di vie parallele, ridisegnando il flusso informativo. Nel genoma umano, il repertorio delle chinasi codificanti è dell’ordine di qualche centinaio di geni, corrispondente a una piccola ma strategica frazione dell’intero gene set proteico; in una cellula di mammifero coesistono molte centinaia di chinasi diverse. Molte proteine segnale presentano multipli siti fosforilabili, ciascuno dedicato a chinasi differenti: la combinazione degli stati fosforilativi agisce come dispositivo logico che integra ingressi multipli in un’unica uscita. Tre meccanismi frequenti di integrazione includono:
- somma e sottrazione di segnali via fosforilazioni su siti distinti, con bilancio tra chinasi e fosfatasi;
- feed‑forward e feedback che generano soglie, memoria e oscillazioni, influenzando sensibilità e dinamica;
- cooperatività nella fosforilazione multi‑sito, che può produrre risposte ultrasensibili con comportamento tipo interruttore.
La cooperatività può essere descritta da una funzione tipo Hill, in cui la frazione di molecole attivate A dipende dalla concentrazione di segnale S:
\[ A(S) = \frac{S^{n}}{K^{n} + S^{n}}, \]
dove n>1 riflette l’ultrasensibilità emergente da più eventi di fosforilazione coordinati. In questo modo, reti chinasi e adattatori proteici si comportano come circuiti computazionali distribuiti, capaci di filtrare rumore, eseguire decisioni binarie o risposte graduali, e veicolare l’informazione su molteplici bersagli a valle. La mappatura di queste reti è in rapido avanzamento: nuove componenti e collegamenti vengono costantemente identificati, così come vie finora sconosciute. Sebbene il sequenziamento dei genomi abbia fornito elenchi completi di attori della trasduzione, la sfida cruciale rimane capire come, nel contesto cellulare nativo, tali elementi cooperino nel tempo e nello spazio per trasformare una combinazione mutevole di segnali ambientali in comportamenti che massimizzino adattamento e sopravvivenza.


