


La scienza trae beneficio da un approccio cooperativo e da punti di vista differenti
Definizione

Fra le molte tipologie cellulari osservabili al microscopio, i batteri rappresentano un modello di organizzazione minima della vita. Una cellula procariotica non possiede compartimenti delimitati da membrane, fatta eccezione per la membrana plasmatica; gli unici complessi macromolecolari ubiqui sono i ribosomi. Il materiale genetico non è racchiuso in un nucleo, ma organizzato in una regione detta nucleoide. La presenza o l’assenza di un nucleo funge da criterio fondamentale per distinguere due grandi gruppi di organismi: gli eucarioti, le cui cellule hanno un nucleo vero e proprio, e i procarioti, privi di nucleo e di organuli membranosi interni.
La morfologia procariotica è varia ma ricorrente in poche forme principali: sferiche (cocchi), a bastoncello (bacilli), spiralate o elicoidali (spirilli e spirochete), cui si aggiungono occasionalmente filamenti, forme vibrioniche o cellule appiattite (Figura 01.04-01). Le dimensioni sono in genere ridotte, spesso tra 0,5 e 5 micrometri, benché siano noti esempi macroscopici eccezionali decine o centinaia di volte più grandi. La cellula è avvolta dalla membrana plasmatica e, nella maggior parte dei casi, da un involucro esterno di protezione, la parete cellulare, con architettura chimica distinta nei batteri Gram-positivi e Gram-negativi. Alcune specie presentano ulteriori strati superficiali proteici (S-layer) o capsule polisaccaridiche che contribuiscono a resistenza e adesione.
All’interno, il citoplasma si presenta come una matrice granulare priva di compartimentalizzazione visibile al microscopio elettronico convenzionale (Figura 01.04-02). Il DNA cromosomico, generalmente circolare, è concentrato nel nucleoide; plasmidi autonomi possono ospitare geni accessori, spesso coinvolti in resistenze o vie metaboliche specializzate. Benché privi di organuli, i procarioti possiedono un citoscheletro proteico (ad esempio FtsZ, MreB, CreS) che governa divisione, forma e segregazione del DNA. Inclusioni di riserva (glicogeno, granuli di zolfo o polifosfato, vescicole gassose) consentono una gestione efficiente delle risorse.
La riproduzione avviene prevalentemente per scissione binaria, con tempi generazionali molto brevi in condizioni favorevoli. Una descrizione quantitativa della crescita esponenziale è data da \(N(t) = N_0 \cdot 2^{t/\tau}\), dove \(N_0\) è il numero iniziale di cellule, \(t\) il tempo e \(\tau\) il tempo di raddoppio. Per esempio, con \(\tau = 18\) minuti, dopo 9 ore si ottiene \(N(9\ \text{h}) = N_0 \cdot 2^{30}\), ossia oltre un miliardo di cellule a partire da una sola. In natura, tuttavia, la crescita è modulata da limiti di nutrienti, competizione, predazione e stress ambientali; alcune specie formano endospore altamente resistenti in risposta a condizioni avverse.
Nonostante la riproduzione asessuata, l’evoluzione procariotica è accelerata dalla combinazione di rapidi tempi di generazione, elevata numerosità e trasferimento orizzontale di geni. Lo scambio di materiale genetico avviene tramite più vie:
- trasformazione, con captazione di DNA libero dall’ambiente;
- coniugazione, mediata da contatto fisico e passaggio di plasmidi o segmenti cromosomici attraverso pili;
- trasduzione, veicolata da batteriofagi che trasferiscono frammenti di DNA tra cellule.
Questi processi, insieme a mutazioni e riarrangiamenti, consentono l’acquisizione rapida di nuovi tratti, come l’utilizzo di substrati insoliti o la resistenza a molecole antimicrobiche. Sistemi immunitari adattativi, quali CRISPR-Cas, mitigano l’ingresso di elementi genetici indesiderati pur non impedendo l’innovazione evolutiva.
Image Gallery
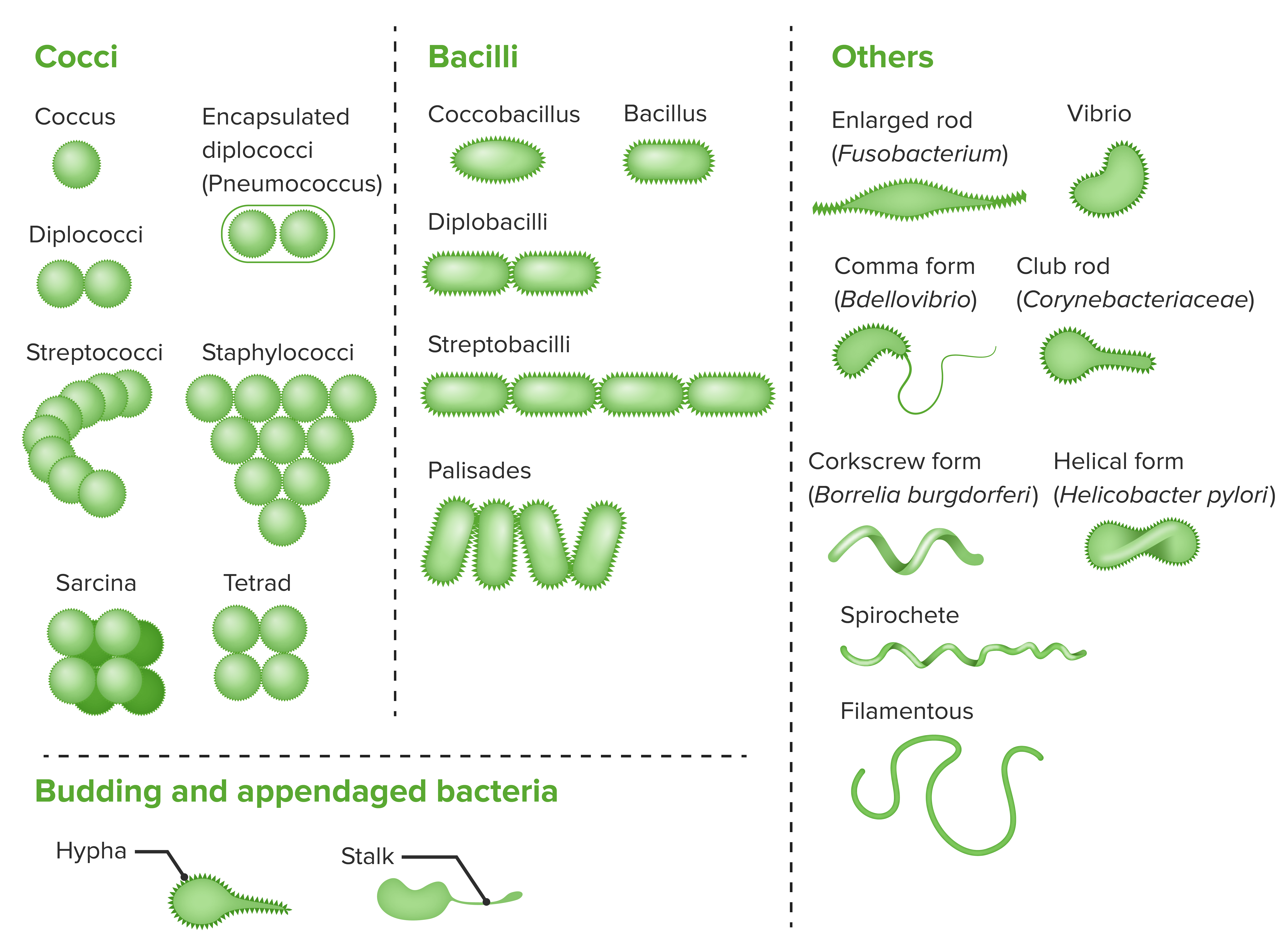
Image Gallery

La maggior parte dei procarioti vive come cellule isolate, ma molte specie formano colonie, catene o complessi biofilm con differenziazione funzionale. Pur apparendo semplici nella forma, i procarioti sono incomparabili per diversità biochimica e capacità metaboliche. Essi colonizzano quasi ogni ecosistema: suoli, oceani, acque dolci, superfici rocciose, pori minerali, il microbiota degli animali e l’interno di cellule eucariotiche. Numericamente dominano la biosfera, con stime globali dell’ordine di 10^30 cellule.
La flessibilità respiratoria è ampia: si trovano specie aerobie obbligate, anaerobie strette sensibili all’ossigeno, anaerobi facoltativi, microaerofili e organismi capaci di respirazioni alternative usando come accettori finali composti quali nitrati, solfati o ossidi di metalli. Questa varietà si accompagna a strategie energetiche differenti:
- fotoautotrofi, che impiegano l’energia luminosa per fissare il carbonio a partire da CO₂ (Figura 01.04-03);
- chemioautotrofi (chemiolitotrofi), capaci di trarre energia dall’ossidazione di composti inorganici ridotti, come H₂, NH₃, Nitrito, Fe²⁺ o H₂S, sintetizzando sostanza organica da CO₂ (Figura 01.04-04);
- eterotrofi, che degradano composti organici complessi, inclusi materiali recalcitranti come lignina e idrocarburi.
Queste attività collocano i procarioti al centro dei cicli biogeochimici. La fissazione dell’azoto atmosferico (N₂) in ammoniaca, effettuata da diazotrofi specializzati, rende biodisponibile un elemento essenziale che le piante, da sole, non sanno sottrarre all’atmosfera. Altri procarioti ossidano ammoniaca a nitrito e nitrato (nitrificazione) o riducono nitrato ad azoto (denitrificazione), chiudendo il ciclo. Nel ciclo del carbonio, batteri e archei partecipano sia alla produzione primaria sia alla decomposizione e al riciclo della materia organica.
La storia evolutiva delle cellule eucariotiche è profondamente intrecciata con i procarioti. Sulla base di molteplici evidenze genetiche e biochimiche, i mitocondri derivano da un’antica endosimbiosi con batteri aerobi, mentre i cloroplasti hanno origine da cianobatteri fotosintetici inglobati da un antenato delle piante e delle alghe. In questo senso, il metabolismo ossidativo e la fotosintesi delle eucariote moderne riflettono capacità procariotiche acquisite e stabilizzate nel tempo.
Image Gallery

Image Gallery

La tassonomia classica considerava i procarioti come un unico raggruppamento. Gli studi di sistematica molecolare, in particolare le analisi dell’RNA ribosomiale 16S/18S, hanno rivelato invece una profonda dicotomia che ha portato al riconoscimento di due domini distinti: Bacteria e Archaea. Queste linee evolutive si sarebbero separate da un antenato comune più di 3,5 miliardi di anni fa e, a livello di relazioni filogenetiche, risultano tra loro distanti quanto lo sono rispetto alle eucariote.
Il dominio Bacteria comprende la maggior parte dei procarioti incontrati nella vita quotidiana: specie del suolo e delle acque, mutualisti e commensali, ma anche numerosi patogeni. Gli Archaea condividono con i batteri molte caratteristiche morfologiche e organizzative, ma presentano tratti molecolari peculiari. Essi popolano sia ambienti “ordinari” sia nicchie estreme: saline concentrate, sorgenti idrotermali, sedimenti anossici di acque profonde, deserti freddi e laghi subglaciali antartici, oltre al rumine dei ruminanti, dove metanogeni archeali degradano composti organici producendo metano.
Le differenze tra i due domini, invisibili al semplice esame al microscopio, emergono nitidamente sul piano chimico e genetico:
- involucro cellulare, con peptidoglicano tipico dei batteri e analoghi privi di acido muramico (pseudomureina o S-layer) negli archei;
- membrane con lipidi a legame estere e catene aciliche nei batteri, contro lipidi a legame etere con catene isoprenoidi negli archei; in questi ultimi possono formarsi anche monostrati stabili ad alte temperature;
- macchine dell’informazione genetica (RNA polimerasi, fattori di trascrizione, ribosomi) che negli archei somigliano più da vicino a quelle eucariotiche; la sensibilità differenziale ad antibiotici ribosomiali ne è una conseguenza pratica;
- vie metaboliche distintive, tra cui la metanogenesi esclusiva degli archei e ampie capacità fotosintetiche tra i batteri, in particolare nei cianobatteri.
Molti habitat estremi in cui prosperano gli archei richiamano condizioni plausibili della Terra primordiale, quando l’atmosfera era povera di ossigeno e le pressioni selettive erano intense. La comprensione di batteri e archei, pertanto, non solo chiarisce l’ecologia contemporanea del pianeta, ma illumina anche passaggi cruciali dell’evoluzione della vita.


