Gli organismi eucarioti pluricellulari presentano notevoli quantità di DNA non codificante e numerose famiglie multigeniche
TOPICS
Definizione

La comprensione moderna del ciclo fondamentale della riproduzione sessuata affonda le radici in osservazioni compiute nel 1888: si notò che le uova fecondate di un nematode parassita possedevano quattro cromosomi, mentre i gameti dello stesso organismo (spermatozoi e uova non fecondate) ne mostravano soltanto due. Da tale dato scaturì la conclusione che i gameti debbano originare tramite una divisione cellulare “riduttiva”, capace di dimezzare con precisione il numero dei cromosomi. A questa divisione fu attribuito il termine meiosi (dal greco meiosis, “diminuzione”). Le indagini di Theodor Boveri, estese ai vermi e ad altre specie, resero evidente che il comportamento dei cromosomi — allora identificati come corpuscoli colorabili di funzione ignota — rifletteva un principio cardine dell’ereditarietà: i due genitori contribuiscono in misura equivalente al patrimonio genetico della progenie, nonostante l’asimmetria dimensionale tra uovo e spermatozoo. Questi risultati, corroborati in seguito dalla teoria cromosomica dell’ereditarietà proposta da Boveri e Sutton, costituirono una prova decisiva del ruolo dei cromosomi quali portatori dell’informazione genetica. Lo studio della riproduzione sessuata e della meiosi è quindi centrale nella storia e nella pratica della biologia cellulare. Adottando la prospettiva della biologia cellulare contemporanea, l’attenzione si focalizza sulla coordinazione temporale e spaziale dei cromosomi durante la meiosi: gli omologhi si riconoscono e si appaiano (sinapsi), scambiano segmenti tramite ricombinazione e quindi si separano in modo ordinato, rimescolando alleli di origine materna e paterna e generando nuove combinazioni. Eventuali deviazioni da tale programma possono produrre errori di segregazione. Il processo si completa con la fecondazione, in cui due gameti aploidi si fondono ripristinando la diploidia e dando origine a un individuo geneticamente distinto da entrambi i genitori:
- appaiamento degli omologhi e formazione della sinapsi, spesso mediata da un complesso proteico specifico;
- ricombinazione con formazione di chiasmi, essenziale per la coesione funzionale e la corretta disgiunzione degli omologhi;
- segregazione sequenziale degli omologhi e, successivamente, delle cromatidi sorelle;
- fusione gametica e ripristino del numero diploide di cromosomi.
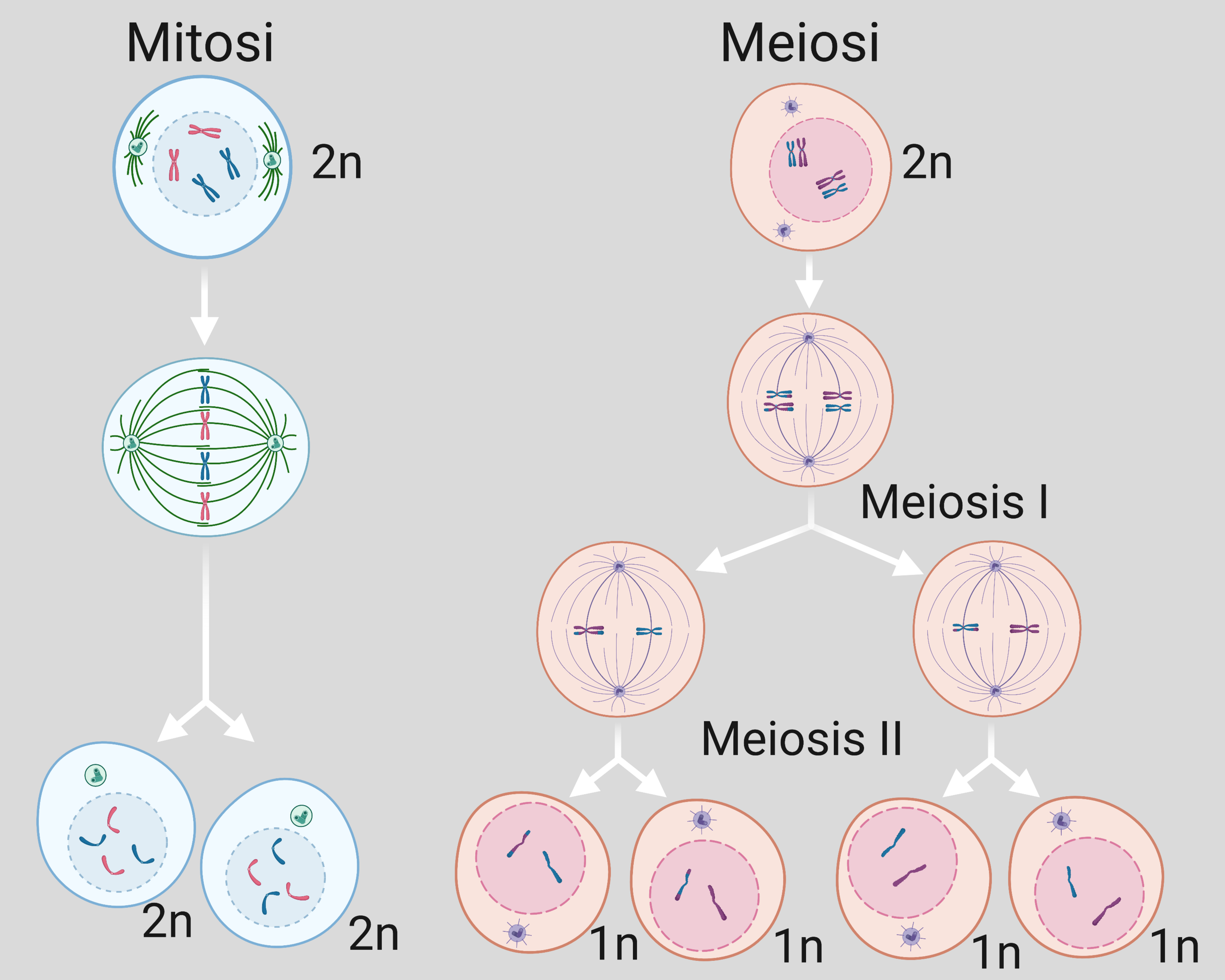
Nella mitosi la cellula replica accuratamente l’intero corredo cromosomico prima della divisione, così che ciascuna delle due cellule figlie riceva una copia completa dei cromosomi di origine materna e una di origine paterna. Anche la meiosi si apre con una fase di replicazione del DNA: tuttavia, la riduzione del numero dei cromosomi non dipende dall’omissione della fase S, bensì dalla presenza di due divisioni cellulari successive che seguono un’unica replicazione iniziale (Figura 07.12-01). La duplicazione produce per ciascun cromosoma una coppia di cromatidi sorelle coese; tale coesione, insieme all’appaiamento tra omologhi, è necessaria per la corretta disgiunzione nelle fasi successive. La meiosi prende avvio in cellule diploidi specializzate della linea germinale localizzate nelle gonadi. Come le cellule somatiche, esse possiedono due versioni di ciascun cromosoma: un omologo di origine paterna e uno di origine materna. Dopo la replicazione, i cromosomi omologhi si riconoscono e si appaiano strettamente (sinapsi). Questa fase, peculiare della meiosi, stabilisce l’architettura necessaria affinché, nelle due divisioni che seguono, ogni gamete riceva un corredo aploide completo. Le due divisioni sono distinte per natura e risultato: la divisione meiotica I (meiosi I) è “riduzionale”, perché separa i cromosomi omologhi, mantenendo unite le cromatidi sorelle; la divisione meiotica II (meiosi II) è “equazionale”, analoga alla mitosi, e separa le cromatidi sorelle. Al termine del processo si ottengono quattro cellule aploidi geneticamente differenti, ognuna con metà del numero di cromosomi della cellula germinale di partenza. L’allocazione degli omologhi ai gameti è casuale: l’orientamento indipendente delle coppie di omologhi in metafase I genera, in assenza di ricombinazione, \(2^n\) assortimenti possibili (dove \(n\) è il numero di coppie di cromosomi); per esempio, con \(n=4\) sono possibili 16 combinazioni distinte. La ricombinazione incrementa ulteriormente la variabilità genetica, rimescolando alleli lungo i cromosomi. La meiosi e la mitosi differiscono dunque per logica e finalità. La prima produce quattro cellule aploidi non identiche, funzionali alla riproduzione sessuata; la seconda genera due cellule figlie diploidi tra loro geneticamente uguali. Tali distinzioni, possono essere riassunte come segue:
- schema replicativo: una sola replicazione del DNA che precede due divisioni nella meiosi, una replicazione seguita da una singola divisione nella mitosi;
- oggetti della segregazione: omologhi in meiosi I e cromatidi sorelle in meiosi II, cromatidi sorelle in mitosi;
- ricombinazione: presente tra omologhi in profase I con formazione di chiasmi, assente nella mitosi;
- esito ploidico: quattro cellule aploidi nella meiosi, due cellule diploidi nella mitosi;
- funzione biologica: generazione di diversità genetica e produzione di gameti nella meiosi, crescita e rinnovamento tissutale nella mitosi.
In sintesi, la sequenza “replicazione unica → due divisioni” è la chiave della riduzione cromosomica. L’appaiamento degli omologhi, la formazione dei chiasmi e la segregazione ordinata consentono di ottenere gameti aploidi competenti, mentre la fecondazione, fondendo due genomi aploidi, ripristina la condizione diploide con un nuovo assetto di alleli rispetto ai genitori.
Image Gallery
Prima di intraprendere sia la mitosi sia la meiosi, una cellula replica l’intero patrimonio cromosomico, generando per ciascun cromosoma una coppia di cromatidi fratelli mantenuti coesi lungo tutta la loro estensione. Nella mitosi, i cromosomi duplicati si dispongono senza un ordine legato all’origine parentale sulla piastra metafasica e, al momento opportuno, i cromatidi fratelli si separano verso i due poli, producendo due cellule figlie geneticamente equivalenti. In meiosi, al contrario, lo scopo di ridurre il numero cromosomico da \(2n\) a \(n\) introduce esigenze addizionali per il sistema di divisione: la cellula germinale deve distinguere e coordinare i cromosomi omologhi di origine materna e paterna, così che, al termine del processo, ciascuna delle quattro cellule aploidi riceva un solo cromatide per ogni omologo. Per soddisfare tale requisito, la meiosi si avvia con un lungo e complesso stadio di accoppiamento, durante la profase meiotica (profase I), in cui le coppie di omologhi replicati si riconoscono e si appaiano prima dell’allineamento sulla piastra metafasica . L’unità strutturale che ne deriva è il bivalente (o tetrade), costituito da quattro cromatidi fratelli che restano associati fino alla separazione. Nella prima divisione meiotica si separano gli omologhi materni e paterni; nella seconda divisione si scindono i cromatidi fratelli di ciascun cromosoma. Le modalità con cui gli omologhi si riconoscono non sono del tutto chiarite. In molte specie l’avvio del contatto dipende dall’interazione tra sequenze di DNA corrispondenti distribuite lungo i cromosomi; in aggiunta, movimenti nucleari guidati dai telomeri e la riorganizzazione dell’impalcatura proteica “assiale” dei cromosomi favoriscono l’incontro tra partner omologhi, inclusi i cromosomi sessuali quando presenti. Una volta stabilito, l’appaiamento è altamente stabile e può persistere per l’intera profase I, fase che in alcuni organismi dura mesi o anni. Durante questa transizione, l’architettura dei cromosomi duplicati è governata da complessi coesivi specifici della meiosi che mantengono uniti i cromatidi fratelli, contribuendo alla corretta costruzione dei bivalenti (Figura 07.12-02).
La descrizione puramente “meccanica” della prima divisione meiotica trascura un aspetto cruciale: quasi tutti gli organismi a riproduzione sessuale associano all’appaiamento una ricombinazione omologa estesa, ossia uno scambio di informazioni genetiche tra sequenze nucleotidiche uguali o molto simili. La ricombinazione omologa, impiegata anche come via di riparazione dei danni al DNA, in meiosi coinvolge cromatidi non fratelli all’interno di ciascun bivalente e culmina nello scambio fisico di segmenti cromosomici, un processo denominato crossing over (Figura 07.12-03). Il crossing over è avviato dalla formazione di rotture a doppio filamento nel DNA e procede attraverso fasi coordinate di elaborazione delle estremità, invasione dell’elica omologa, sintesi guidata da stampo e risoluzione delle giunzioni di Holliday. In seguito, porzioni corrispondenti dei cromatidi non fratelli vengono scambiate, generando nuovi assetti allelici. Tale sequenza di eventi è resa preciso dall’allineamento su scala nanometrica degli assi cromosomici assicurato dal complesso sinaptonemico, una impalcatura proteica che connette strettamente i due omologhi replicati, stabilizza i bivalenti e consente lo scambio con accuratezza anche in regioni molto ridotte (Figura 07.12-04). Ogni cromatidio può partecipare a uno o più eventi di crossing over con il cromatidio dell’omologo, benché l’organizzazione dei cross-over tenda a risultare spazialmente regolata lungo il cromosoma. Verso la fine della profase I, il complesso sinaptonemico si disassembla ma i siti ricombinanti maturi persistono come chiasmi, strutture citologiche a forma di incrocio visibili al microscopio (Figura 07.12-05). I chiasmi mantengono uniti tra loro gli omologhi non fratelli e trasformano gli scambi molecolari in collegamenti meccanicamente funzionali. Spesso ciascun bivalente presenta molteplici chiasmi, segno di più eventi di crossing over lungo lo stesso paio di omologhi (Figura 07.12-05). Oltre a generare variabilità genetica nei gameti, i crossing over svolgono un ruolo strutturale essenziale: i chiasmi, insieme alla coesione tra cromatidi fratelli, conferiscono ai bivalenti la continuità meccanica necessaria per resistere alle forze del fuso e per posizionarsi correttamente, garantendo così l’ordinata separazione degli omologhi nella prima divisione meiotica (Figura 07.12-06):
- innesco della ricombinazione con rotture a doppio filamento e resecazione delle estremità 5′;
- invasione dell’elica omologa e formazione del D-loop con appaiamento a elevata fedeltà tra sequenze corrispondenti;
- sintesi di DNA e stabilizzazione delle giunzioni ricombinanti;
- risoluzione verso prodotti crossing over o non-crossing over, con i primi che si manifestano citologicamente come chiasmi.
Image Gallery
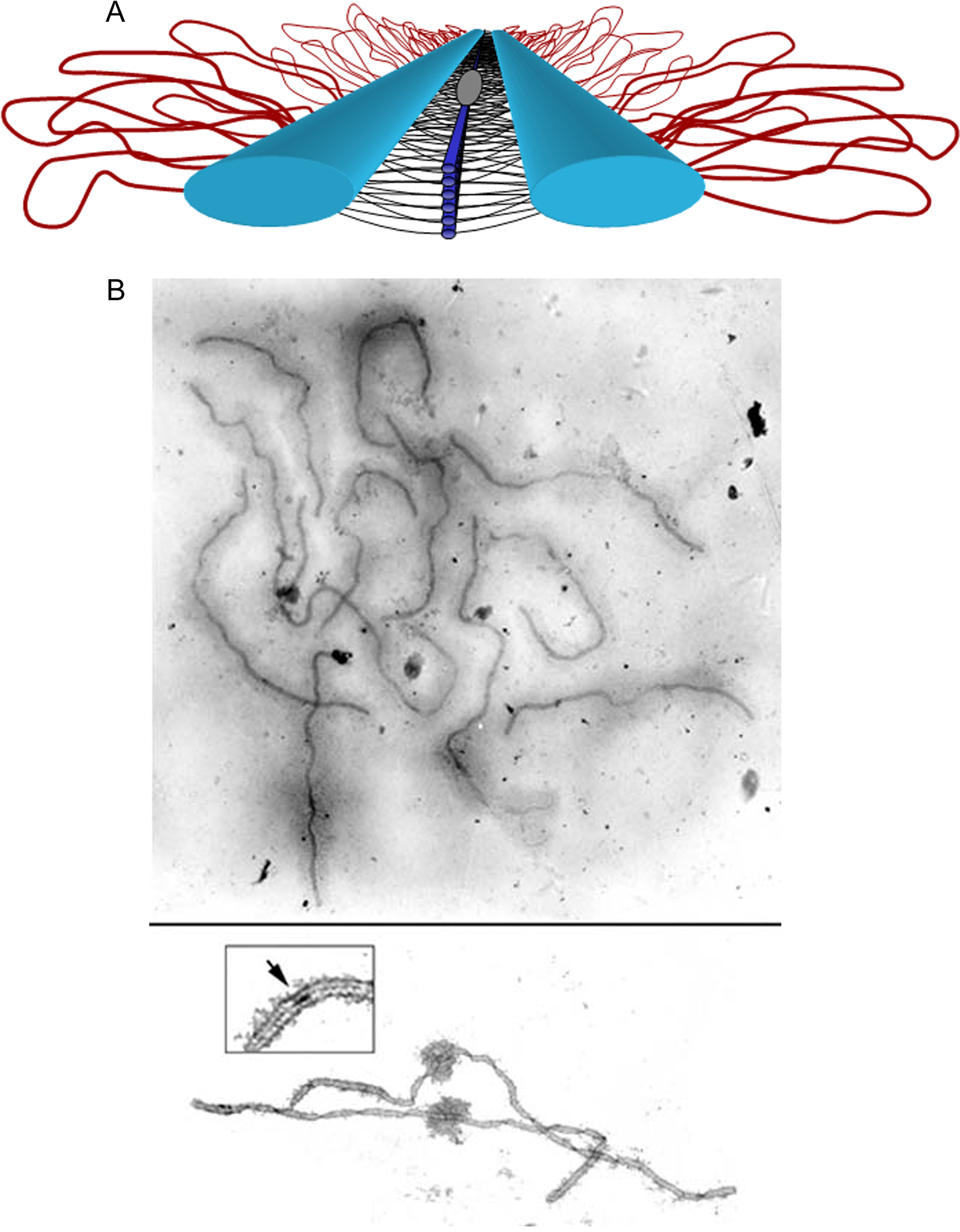
Image Gallery
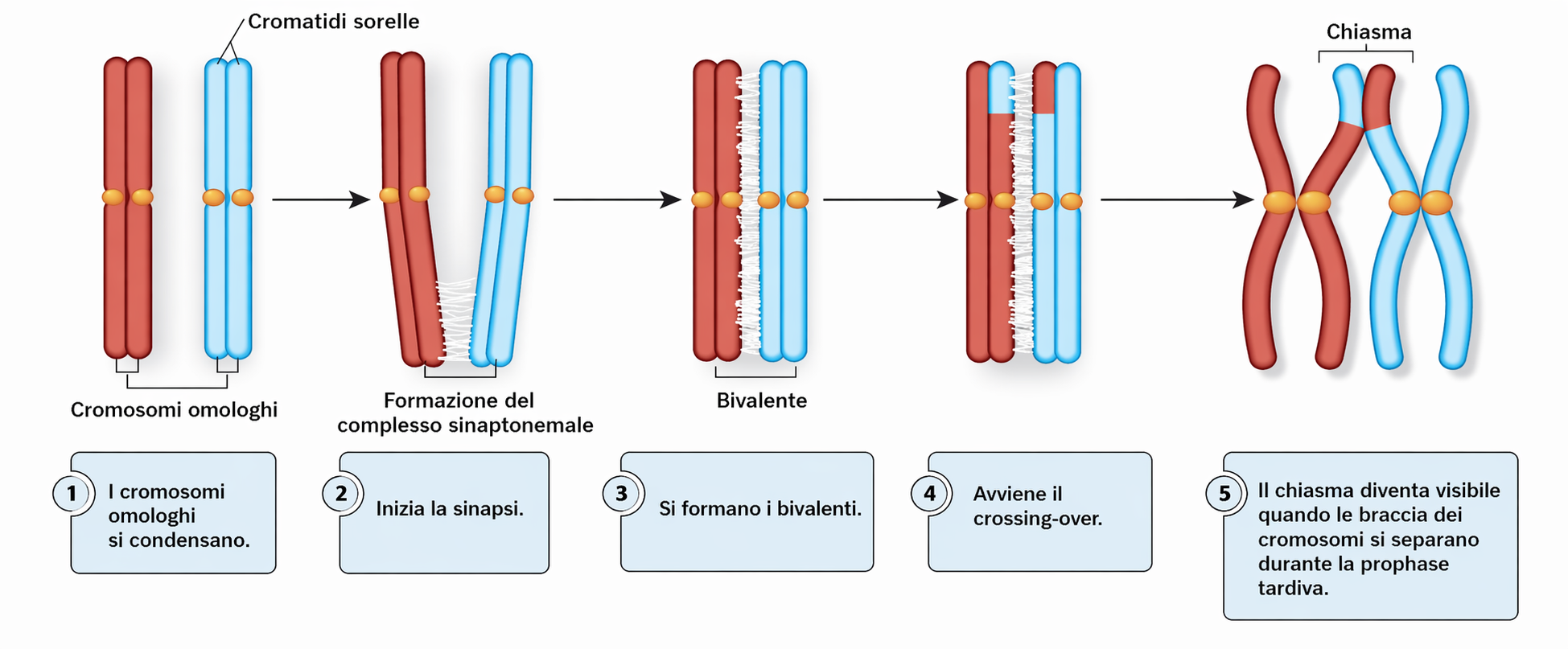
Image Gallery
Image Gallery
Nella maggior parte degli organismi, la ricombinazione meiotica è indispensabile per la corretta separazione dei due omologhi replicati in anafase I. I chiasmi derivanti dagli scambi, in cooperazione con la coesione tra cromatidi fratelli, mantengono uniti i partner materno e paterno finché il fuso non esercita trazione opposta sui bivalenti, stabilizzandone l’allineamento sulla piastra metafasica e permettendo il controllo della tensione generata dai microtubuli (Figura 07.12-07). Il legame tra gli omologhi deriva da due contributi complementari: i chiasmi, che collegano cromatidi non fratelli, e le proteine coesine che tengono appaiati i cromatidi fratelli lungo i bracci dei cromosomi fino alla meiosi I (Figura 07.12-04). All’inizio dell’anafase I, l’attivazione della separasi determina la rimozione selettiva delle coesine lungo i bracci, permettendo il disancoraggio dei chiasmi e la separazione degli omologhi verso poli opposti (Figura 07.12-07). La coesione centromerica, invece, è protetta fino alla meiosi II da specifici fattori, così che i cromatidi fratelli rimangano congiunti e si orientino congiuntamente verso lo stesso polo nella prima divisione, per poi segregare tra loro nella seconda:
- chiasmi come collegamenti meccanici tra omologhi che forniscono resistenza alla trazione del fuso;
- coesione dei bracci che mantiene l’integrità dei bivalenti fino alla risoluzione in anafase I;
- protezione della coesione centromerica, che preserva l’unità dei cromatidi fratelli fino all’anafase II;
- orientamento congiunto dei cinetocori dei cromatidi fratelli in meiosi I, essenziale per segregare gli omologhi e non i fratelli.
La perdita di chiasmi adeguati o un difetto nella dinamica della coesione compromette la separazione degli omologhi, favorendo eventi di non-disgiunzione e la formazione di gameti aneuploidi. L’integrazione tra appaiamento omologo, ricombinazione e controllo della coesione costituisce quindi la base sia della riduzione cromosomica sia dell’accuratezza della segregazione meiotica.
Image Gallery
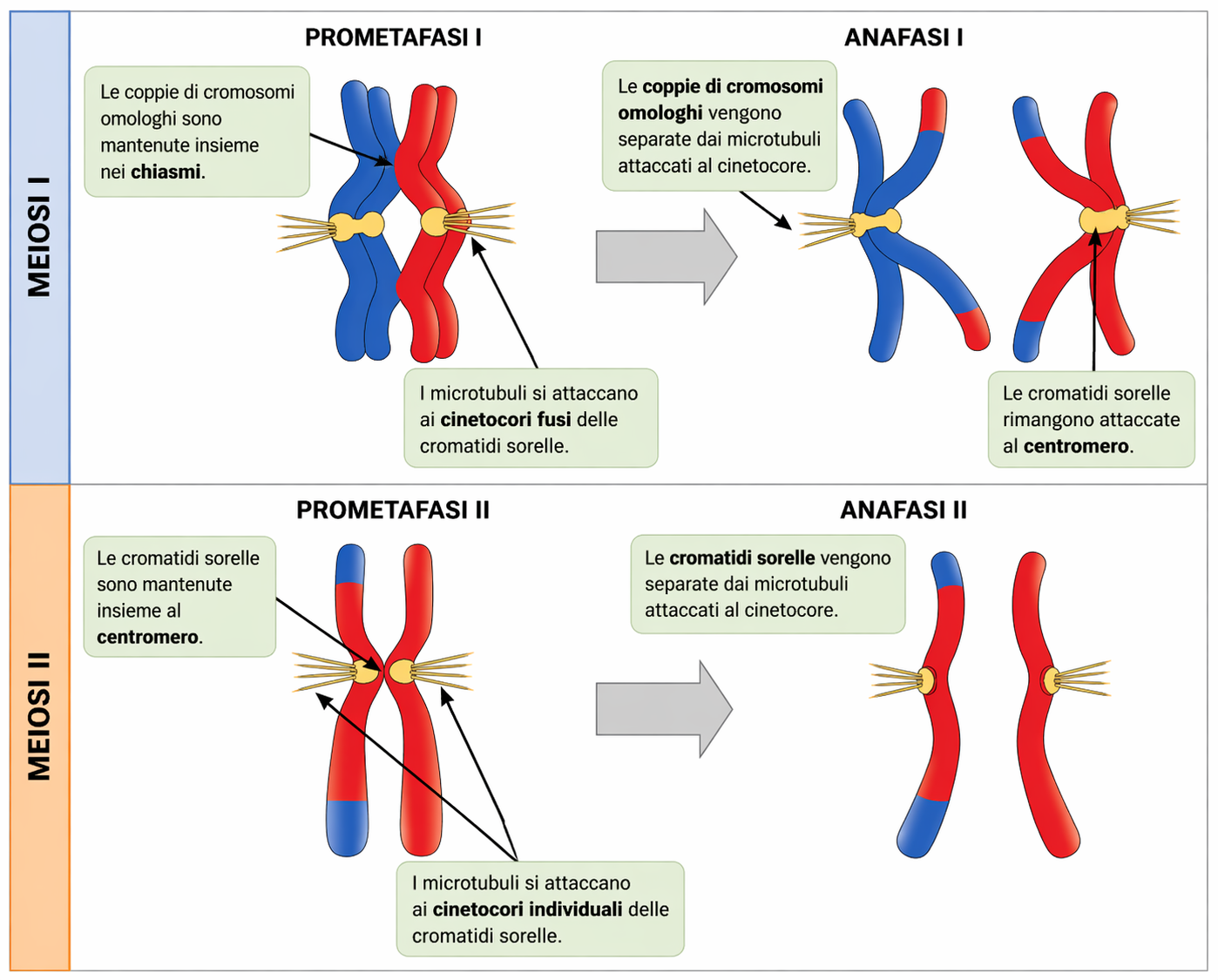
Image Gallery
Image Gallery
Dopo la prima divisione meiotica, le cellule figlie non ripristinano un’interfase completa né duplicano nuovamente il DNA. La seconda divisione, la meiosi II, si avvia rapidamente: si riorganizza un nuovo fuso e i cinetocori dei cromatidi fratelli, che in meiosi I erano co-orientati verso lo stesso polo, acquisiscono ora un’orientazione opposta (bi-orientazione) e si connettono a microtubuli provenienti dai due poli, come accade nella mitosi. Quando si instaura una corretta tensione sui cinetocori, la coesione centromerica residua viene rimossa e i cromatidi fratelli si separano all’anafase II, migrando verso poli opposti e generando cellule figlie che ereditano ciascuna una singola cromatide per cromosoma omologo. Nelle specie animali, la specificità della meiosi si fonda anche sulla regolazione della coesione: la sottounità meiotica delle coesine (ad esempio Rec8 nei modelli sperimentali) è protetta al centromero in meiosi I da proteine “tutrici” (come Shugoshin), così che la separazione avvenga solo tra omologhi; tale protezione è rimossa prima della meiosi II, consentendo la scissione della coesione centromerica da parte della separasi e la separazione dei cromatidi fratelli. Sebbene l’architettura del fuso e la dinamica dei microtubuli ricordino la mitosi, va sottolineato che la meiosi II opera su un assetto cromosomico aploide, con la sola coppia di cromatidi fratelli per ciascun tipo di cromosoma, e produce quindi quattro cellule aploidi complessive con combinazioni genetiche originali.
Individui nati dagli stessi genitori biologici presentano tra loro differenze genetiche estese, a meno che non derivino dallo stesso zigote. La ragione risiede nel rimescolamento casuale dell’informazione genetica che si realizza già in meiosi I tramite due processi indipendenti e complementari:
- assortimento indipendente dei cromosomi omologhi: i cromosomi di origine materna e paterna, organizzati in bivalenti, si orientano casualmente sul fuso della metafase I; la cattura dei cinetocori da parte dei microtubuli determina quale omologo si dirigerà a ciascun polo, per cui ogni gamete riceve una combinazione stocastica di cromosomi materni e paterni; si veda anche l’orientamento riassunto in (Figura 07.12-07);
- ricombinazione meiotica (crossing-over): all’interno di ciascun bivalente l’appaiamento sinaptinemale favorisce scambi di tratti omologhi tra cromatidi non fratelli; ogni singolo evento di crossing-over genera alle combinazioni nuove di alleli lungo i cromosomi, e in media nelle cellule umane si osservano due o tre scambi per coppia di omologhi, con localizzazione non deterministica lungo i bracci.
Il solo assortimento indipendente, trascurando la ricombinazione, consente teoricamente a un individuo di produrre \(2^n\) tipi di gameti distinti, dove \(n\) è il numero aploide di cromosomi. Nell’uomo, con \(n = 23\), il numero raggiunge \(2^{23} \approx 8,4 \cdot 10^6\). Questo è tuttavia un limite inferiore: gli scambi ricombinativi aumentano enormemente la diversità, perché ogni meiosi produce quattro cromatidi derivanti da ricombinazione differenti tra loro, amplificando il repertorio di aplotipi presenti nei gameti oltre quanto previsto dal solo assortimento casuale. La genesi dei chiasmi non solo genera variabilità, ma garantisce anche la corretta segregazione in meiosi I: i chiasmi, insieme alla coesione tra cromatidi fratelli, creano collegamenti fisici tra gli omologhi che assicurano una tensione bilanciata al fuso. Poiché le posizioni dei crossing-over sono variabili e gli omologhi di ciascuna coppia possono essere scelti in molteplici combinazioni, la variazione risultante nelle cellule aploidi è sostanzialmente inesauribile. Per illustrare l’ampiezza del rimescolamento, si consideri un caso ipotetico: due genitori portatori di alleli diversi per quattro loci su due cromosomi non omologhi possono produrre gameti con tutte le combinazioni possibili degli alleli tra i loci indipendenti, cui si sommano varianti generate da un crossing-over che separi alleli originariamente associati sullo stesso cromosoma. Insieme, questi meccanismi spiegano perché, anche all’interno della stessa famiglia, ciascun figlio presenti un profilo genetico unico pur partendo dallo stesso patrimonio parentale.
Image Gallery
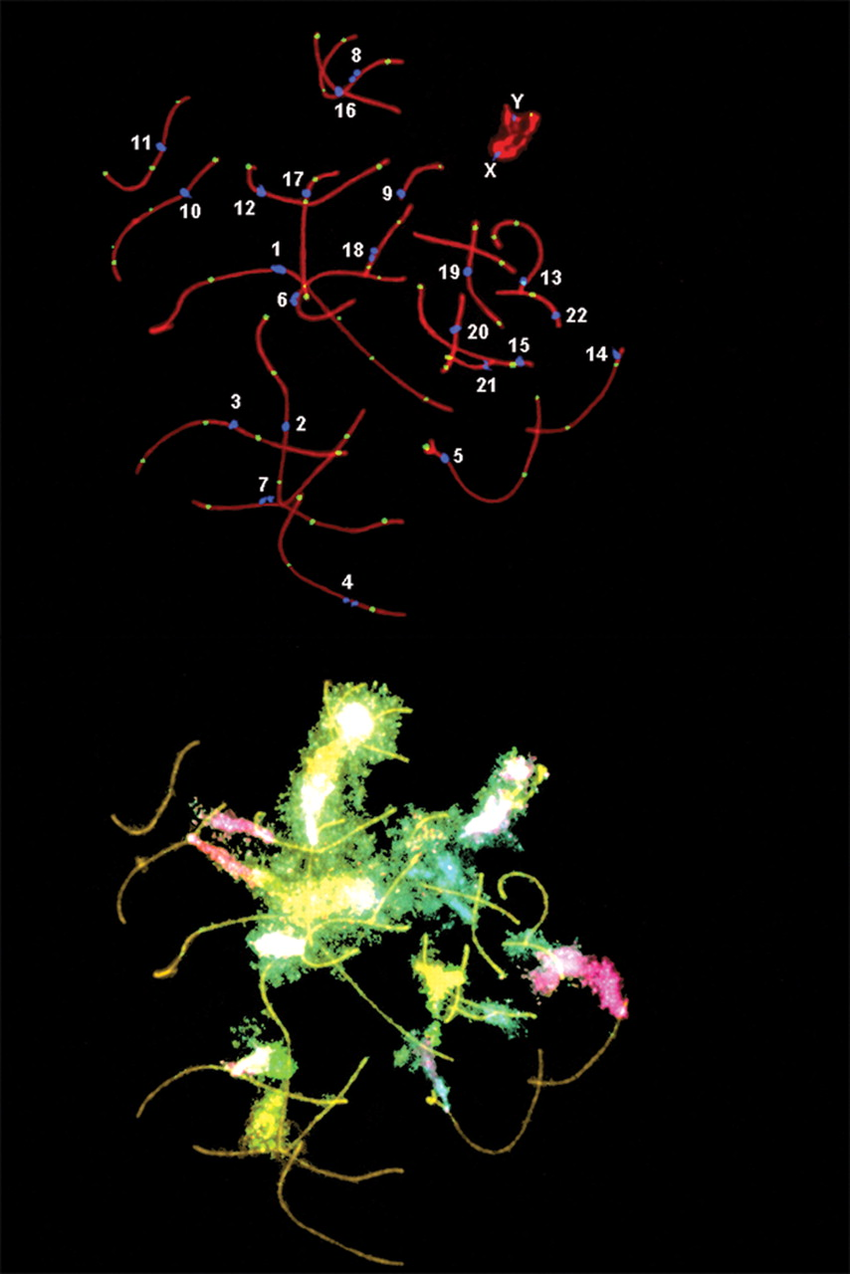
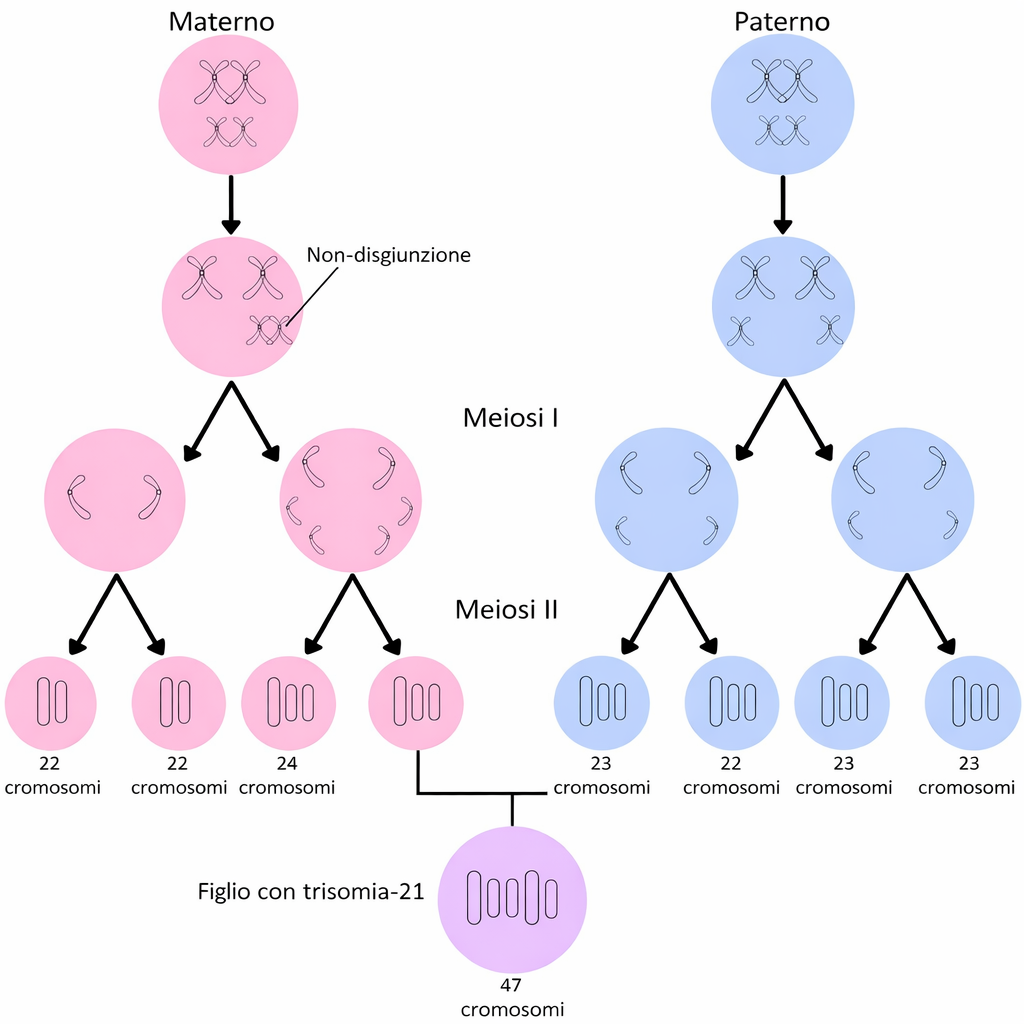
La segregazione cromosomica durante la meiosi è un processo di elevata complessità: nella specie umana, ogni meiosi parte da 23 coppie di omologhi duplicati (92 cromatidi) e deve distribuire in modo accurato una singola copia di ciascun cromosoma in ogni gamete. Non sorprende che si possano verificare errori, anche in presenza di sistemi di controllo stringenti. Un errore ricorrente è la non disgiunzione, ovvero il fallimento nella separazione di omologhi in meiosi I o di cromatidi fratelli in meiosi II. Il risultato sono gameti aneuploidi, privi di un cromosoma o contenenti una copia in eccesso. Dopo la fecondazione, questi gameti originano zigoti con squilibrio cromosomico. Spesso l’embrione così formato non è vitale e va incontro a morte precoce, ma alcune aneuploidie sono compatibili con lo sviluppo. Un esempio rilevante è la trisomia 21: la presenza di tre copie del cromosoma 21, tipicamente dovuta a non disgiunzione in meiosi I del gamete femminile, conduce alla sindrome di Down, caratterizzata da alterazioni dello sviluppo e peculiarità fenotipiche; l’evento è illustrato schematicamente in (Figura 07.12-08). L’incidenza degli errori di segregazione è particolarmente elevata negli ovociti umani, dove si possono osservare frequenze intorno al 10% di meiosi con non disgiunzione e conseguente aneuploidia dell’uovo. Negli spermatozoi l’aneuploidia è meno comune, probabilmente per l’eliminazione preferenziale delle cellule germinali con errori mediante meccanismi di sorveglianza del ciclo cellulare e apoptosi. Tra i fattori predisponenti si annoverano la riduzione dell’integrità della coesione con l’età materna, la localizzazione atipica dei crossing-over (ad esempio troppo prossimi a centromeri o telomeri) e un controllo subottimale del checkpoint di assemblaggio del fuso, che normalmente rileva l’assenza di tensione ai cinetocori e ritarda l’anafase. Oltre alla trisomia 21, altre aneuploidie autosomiche (come trisomia 18 e 13) e dei cromosomi sessuali (ad esempio 45,X o 47,XXY) possono manifestarsi, sebbene con esiti clinici e viabilità differenti. Indipendentemente dal gamete in cui si è verificato l’errore, la non disgiunzione rappresenta una causa maggiore dell’elevata frequenza di aborti spontanei nelle fasi precoci della gestazione.
Image Gallery
La ricostituzione del genoma diploide avviene con la fecondazione, quando i gameti aploidi si fondono per formare uno zigote. Nell’uomo, dei centinaia di milioni di spermatozoi depositati durante il coito, solo una frazione minima raggiunge l’ovidotto, sede tipica dell’incontro con l’ovocita. La navigazione degli spermatozoi è modulata da segnali chimici rilasciati dall’ovocita e dal cumulo ooforo circostante. Al contatto, lo spermatozoo deve attraversare gli strati cellulari di rivestimento, legarsi alla zona pellucida, superarla e infine fondersi con la membrana plasmatica dell’ovocita (Figura 07.12-09). Questo percorso si accompagna a eventi molecolari chiave, tra cui la reazione acrosomiale e l’interazione tra proteine di membrana specifiche dei gameti. Benché numerosi spermatozoi possano aderire all’uovo, normalmente uno soltanto completa la fusione di membrana e rilascia il proprio genoma nel citoplasma ovocitario. Tale unicità è assicurata da meccanismi che impediscono la polispermia. Nella specie umana, l’ingresso del primo spermatozoo induce un’onda di ioni Ca²⁺ nel citoplasma, che promuove l’esocitosi dei granuli corticali; gli enzimi liberati rimodellano la zona pellucida, rendendola refrattaria a ulteriori penetrazioni. La stessa cascata di Ca²⁺ contribuisce ad attivare l’ovocita, concludendo la meiosi II e predisponendo i processi di sviluppo embrionale precoce. La fecondazione può avvenire anche con tecniche assistite: l’iniezione intracitoplasmatica dello spermatozoo (ICSI) supera, quando necessario, difetti di fusione o motilità spermatiche inserendo direttamente un singolo spermatozoo nel citoplasma ovocitario. Indipendentemente dalla modalità, la fecondazione si considera completa quando i due pronuclei aploidi, uno di origine paterna e l’altro materna, migrano, si avvicinano e fondono i loro corredi cromosomici in un unico nucleo diploide. Da questo momento, lo zigote entra in una serie coordinata di divisioni mitotiche e programmi di espressione genica che conducono, attraverso lo sviluppo embrionale, alla formazione dell’organismo adulto.
Image Gallery