


Duplicazione, ristrutturazione e mutazioni del DNA contribuiscono all’evoluzione del genoma
TOPICS
Definizione

Nella riproduzione asessuata, il materiale genetico del genitore viene duplicato e trasmesso senza ricombinazione, producendo discendenti praticamente identici al singolo progenitore. Prima dell’opera di Mendel, diversi naturalisti ipotizzavano che l’ereditarietà negli esseri umani potesse seguire una logica analoga, con caratteri trasmessi come copie intatte da una sola linea parentale (Figura 07.13-01). Negli organismi a riproduzione sessuata, invece, la somiglianza familiare non equivale all’identità. I meccanismi meiotici, in particolare l’assortimento indipendente dei cromosomi omologhi e il crossing-over, riorganizzano gli alleli in nuove combinazioni; la successiva fecondazione casuale unisce gameti differenti, generando una prole che mostra una combinazione unica di tratti derivati da entrambi i genitori, talora con fenotipi non osservati in precedenza. La possibilità di seguire nel tempo caratteri che si conservano o si ricombinano da una generazione alla successiva ha consentito di dedurre regolarità generali dell’ereditarietà negli organismi a riproduzione sessuata. I caratteri più agevoli da analizzare sono quelli con manifestazioni nette e misurabili. In ambito umano, esempi classici includono la percezione del feniltiocarbammide (PTC), che dipende da varianti in geni recettoriali del gusto, oppure sistemi ematologici con fenotipi distinguibili in modo univoco, come i gruppi sanguigni, che si determinano con semplici test di laboratorio (Figura 07.13-02). Tuttavia, la chiarificazione delle leggi generali dell’ereditarietà è stata ottenuta studiando organismi facili da allevare e prolifici, nei quali i caratteri discreti sono facili da controllare sperimentalmente. Gregor Mendel, considerato il fondatore della genetica, scelse i piselli da orto; ma approcci analoghi si applicano a moscerini della frutta, nematodi, topi e numerose piante coltivate. Le regole fondamentali ricavate da questi sistemi valgono per tutti gli organismi che si riproducono sessualmente, comprese le specie umane. Nei paragrafi seguenti viene ricostruita la logica dell’ereditarietà nei viventi a riproduzione sessuata. La segregazione degli omologhi in meiosi e la loro unione casuale mediante fecondazione spiegano in termini cromosomici i risultati sperimentali e le regole dedotte a partire dai dati. Prima, però, è utile ripercorrere come Mendel, oltre 150 anni fa, riuscì a identificare tali regolarità coltivando piante di pisello nell’orto del suo monastero.
Image Gallery
Image Gallery

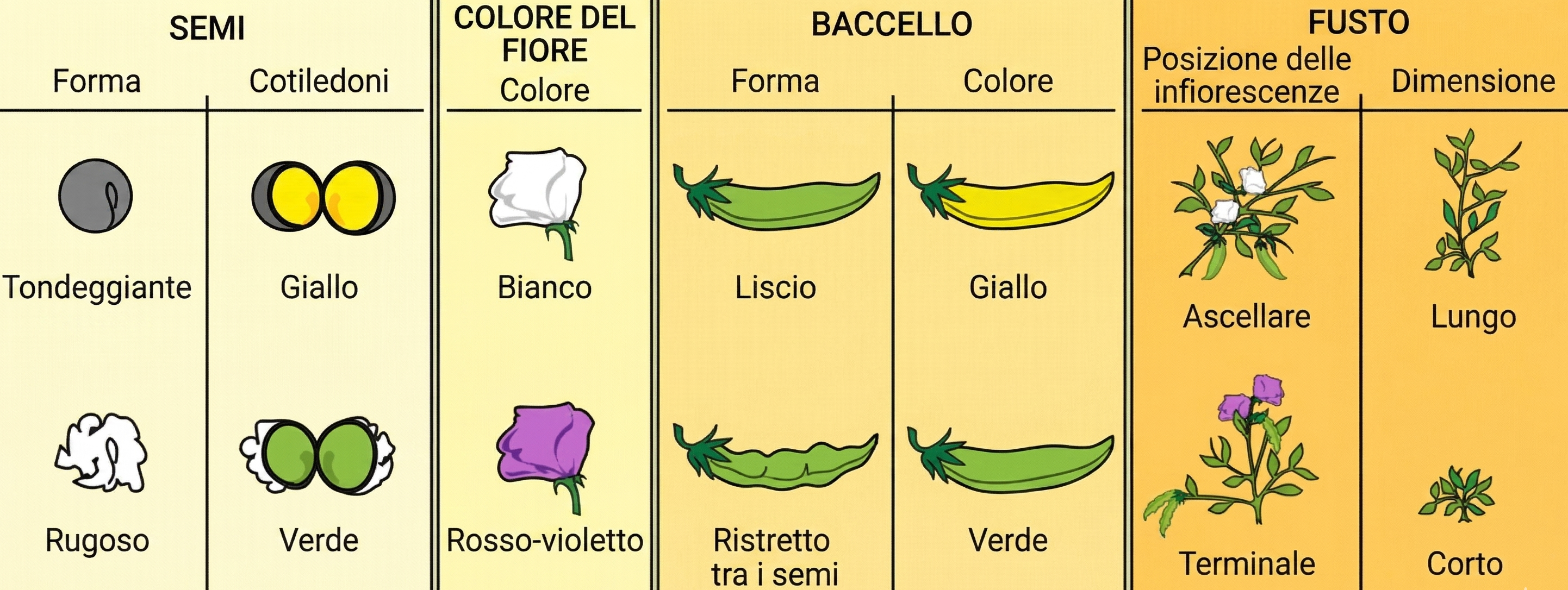
Mendel scelse i piselli perché consentivano di lavorare con numeri elevati in spazi ridotti, garantivano cicli riproduttivi brevi e, soprattutto, permettevano il controllo degli incroci. Egli trasferiva manualmente il polline da una pianta a un’altra, controllando con precisione la parentela della progenie ottenuta. Un ulteriore vantaggio era la disponibilità di molte varietà, ciascuna con tratti discreti e facilmente riconoscibili: ad esempio fiori di colore viola o bianco, o semi con tegumento liscio oppure rugoso. Mendel selezionò sette caratteri, ciascuno espresso in due forme alternative ben distinte e poco influenzate da variabili ambientali: la netta distinguibilità dei fenotipi era cruciale per una valutazione non ambigua degli esiti (Figura 07.13-03):
- gli incroci potevano essere diretti e ripetuti in modo controllato;
- le piante producevano numerosa progenie, utile per analisi statistiche;
- le linee “a riproduzione costante” (linee pure) erano disponibili e facilmente mantenibili;
- i caratteri scelti mostravano categorie fenotipiche non sovrapponibili.
Image Gallery
La strategia sperimentale di Mendel era essenziale e rigorosa. Egli iniziò con linee pure, cioè popolazioni che, quando autosseminale, generavano discendenti uniformi per il tratto d’interesse. L’uso continuativo di linee di riferimento gli consentiva di rilevare senza ambiguità eventuali cambiamenti nella progenie. A differenza di molti predecessori che incrociavano organismi divergenti per più caratteristiche, complicando l’interpretazione dei risultati, Mendel considerò un carattere alla volta. In un incrocio tipico, fece impollinare reciprocamente linee pure alternative per un dato tratto, ad esempio semi lisci con semi rugosi, e registrò il fenotipo della prima generazione filiale (F1). Gli ibridi F1 risultarono tutti uniformi e somiglianti a uno solo dei due genitori per il carattere in esame. Se l’analisi si fosse fermata alla F1, si sarebbe potuto concludere, erroneamente, che la prole replica in toto uno dei due contributi parentali. Invece, proseguendo, Mendel lasciò che le piante F1 si autoimpollinassero o le incrociò fra loro, esaminando così la generazione F2.
Il comportamento degli ibridi F1 sollevava un interrogativo decisivo: che fine avevano fatto le varianti fenotipiche non osservate? Gli esperimenti successivi mostrarono che il fenotipo assente in F1 ricompare in F2 con proporzione costante: in media, circa tre quarti degli individui F2 manifestavano il fenotipo osservato in F1, e circa un quarto mostrava l’alternativa. Lo stesso schema si ripeté per tutti i caratteri analizzati. Per interpretare questi dati, Mendel propose che i caratteri siano determinati da fattori ereditari discreti, oggi detti geni, presenti in versioni alternative (alleli) che corrispondono alle varianti fenotipiche osservate. Ogni organismo diploide possiede due copie di ciascun gene, una di origine materna e una di origine paterna. Nelle linee pure, i due alleli sono uguali (omozigosi); negli ibridi F1, i due alleli sono differenti (eterozigosi). La scomparsa di una variante in F1 e la sua ricomparsa in F2 si spiegano postulando che, in una coppia allelica, uno dei due alleli sia dominante e l’altro recessivo. In eterozigosi, l’allele dominante determina il fenotipo, mentre l’allele recessivo rimane non espresso ma non scompare dalla discendenza. Nel caso di un incrocio fra eterozigoti per un singolo gene, le probabilità attese seguono la “legge della segregazione”:
\[ \begin{aligned} P(\text{allele A nei gameti}) &= \tfrac{1}{2},\quad P(\text{allele a nei gameti}) = \tfrac{1}{2};\\ P(AA) &= \tfrac{1}{4},\quad P(Aa) = \tfrac{1}{2},\quad P(aa) = \tfrac{1}{4};\\ \text{rapporto genotipico} &= 1:2:1,\quad \text{rapporto fenotipico (con A dominante)} = 3:1. \end{aligned} \]
Da questa impostazione discendono alcune nozioni chiave:
- genotipo: l’insieme degli alleli posseduti da un individuo per uno o più loci;
- fenotipo: l’espressione osservabile del genotipo, modulata dall’ambiente;
- omozigote/eterozigote: condizione di identità o diversità allelica a un locus;
- dominanza/recessività: relazione funzionale tra alleli che determina l’esito fenotipico in eterozigosi.
Una conseguenza importante è che molti alleli presenti in un individuo non sono visibili nel suo fenotipo perché mascherati in eterozigosi o perché dipendenti dal contesto genetico. Considerato che il genoma umano comprende circa 20 000 geni, ciascuno di noi porta numerose varianti che possono non manifestarsi, ma che possono emergere in combinazioni specifiche nelle generazioni successive attraverso la segregazione meiotica e la fecondazione casuale. Infine, la ricombinazione durante la meiosi e l’assortimento indipendente dei cromosomi omologhi generano nuove combinazioni alleliche tra loci distinti, ampliando ulteriormente la variabilità nella progenie e fornendo la base molecolare per le proporzioni fenotipiche osservate da Mendel nei suoi incroci controllati.
L’ipotesi formulata da Mendel, secondo cui ogni individuo possiede due copie per ciascun gene, una di origine materna e una di origine paterna, solleva un interrogativo funzionale: in che modo un genitore trasmette alla discendenza una sola copia per gene? La risposta risiede nel processo di formazione dei gameti, in cui le due copie (alleli) di ciascun gene si separano, così che ogni gamete risulta portatore di un singolo allele per carattere. Questa idea, enunciata come legge della segregazione, trova oggi fondamento nella biologia dei cromosomi: durante la meiosi I i cromosomi omologhi, ciascuno con i propri alleli, si disgiungono e migrano verso poli opposti, assicurando che uova e spermatozoi siano aploidi. Se consideriamo i caratteri classici del pisello, ogni ovocellula e ogni granulo pollinico riceve, in modo mutuamente esclusivo, un allele per il colore del seme (ad esempio giallo oppure verde), uno per la forma (liscio oppure rugoso), uno per il colore del fiore (viola oppure bianco) e così via. La fecondazione, cioè l’unione casuale di un gamete maschile con uno femminile, ripristina nello zigote la coppia di alleli per ciascun carattere. La casualità dell’incontro tra gameti fa sì che la ricomposizione delle coppie alleliche nello zigote sia governata da regole probabilistiche ben definite. La legge della segregazione afferma dunque che i due alleli che determinano un carattere si separano nella formazione dei gameti e si riuniscono a caso nella generazione successiva. Una pianta F1 ibrida per il colore del seme, con genotipo eterozigote, produrrà due classi equiprobabili di gameti, ciascuna portatrice di un allele diverso. In un’autoimpollinazione, le quattro combinazioni alleliche possibili in F2 daranno luogo a un rapporto genotipico 1:2:1 e, quando uno degli alleli è dominante, a un rapporto fenotipico 3:1. In termini probabilistici, per un carattere con dominanza completa si avrà \(P(\text{fenotipo dominante}) = 3/4\) e \(P(\text{fenotipo recessivo}) = 1/4\). La relazione tra comportamento dei geni e comportamento dei cromosomi durante la meiosi chiarisce la base citologica della prima legge: all’anafase I si separano gli omologhi (e con essi gli alleli), mentre all’anafase II si separano le cromatidi sorelle, mantenendo nei gameti un’unica copia per gene. L’unione casuale dei gameti completa il quadro, traducendo la segregazione in previsioni quantitative verificabili:
- segregazione meiotica: i cromosomi omologhi, che portano alleli alternativi, si disgiungono all’anafase I, garantendo un allele per gamete;
- casualità nella fecondazione: l’incontro tra gameti è stocastico, per cui le combinazioni alleliche nello zigote seguono le leggi del calcolo delle probabilità;
- risultato in F2 per un carattere con dominanza completa: rapporto fenotipico atteso 3:1, coerente con le quattro combinazioni alleliche possibili.
Le osservazioni di Mendel sui piselli si estendono ad animali, piante e funghi che si riproducono per via sessuata, perché la meiosi e la fecondazione sono processi conservati. Egli riprodusse i risultati fondamentali anche in mais e fagiolo; in modo analogo, la genetica moderna ha confermato che la segregazione degli alleli vale in organismi così diversi come moscerini della frutta, lieviti e mammiferi (Figura 07.13-04). Per illustrare un tratto umano monogenico a eredità autosomica recessiva, consideriamo l’albinismo oculocutaneo: varianti loss-of-function in geni della via biosintetica della melanina conducono a ridotta o assente pigmentazione di cute, capelli e occhi. Gli individui affetti sono omozigoti per l’allele recessivo, indicato con a, e hanno genotipo aa; i portatori eterozigoti (Aa) sono in genere fenotipicamente normali poiché l’allele A è sufficiente a garantire un livello funzionale di pigmentazione (Figura 07.13-05). Scenari riproduttivi equivalenti, con stime di rischio, illustrano la legge della segregazione nell’uomo e differiscono da quelli esemplificati in precedenza:
- incrocio tra portatore e individuo affetto (Aa × aa): a ogni gravidanza, \(P(\text{aa}) = 1/2\) e \(P(\text{Aa}) = 1/2\); la probabilità che due figli consecutivi siano entrambi affetti è \((1/2)^2 = 1/4\);
- incrocio tra non affetto non portatore e affetto (AA × aa): tutti i figli saranno portatori (Aa) e fenotipicamente non affetti, \(P(\text{Aa}) = 1\);
- incrocio tra due portatori (Aa × Aa): distribuzione genotipica 1:2:1 con \(P(\text{aa}) = 1/4\), \(P(\text{Aa}) = 1/2\), \(P(\text{AA}) = 1/4\); ad esempio, la probabilità che il primogenito sia non affetto e il secondogenito affetto è \((3/4) \times (1/4) = 3/16\).
Le dimensioni ridotte dei nuclei familiari umani fanno sì che i rapporti attesi (per esempio 3:1) raramente emergano con precisione in una singola sibship, in contrasto con i grandi numeri ottenuti da Mendel nei suoi incroci. Per questo, la genetica umana ricorre a insiemi di famiglie o a più generazioni e sintetizza le informazioni mediante alberi genealogici (pedigree), che rappresentano la presenza o l’assenza del carattere in ciascun membro. La (Figura 07.13-06) mostra un pedigree per un’ipoposia autosomica recessiva e mette in evidenza una implicazione applicativa delle leggi di Mendel: le unioni tra consanguinei, come i cugini di primo grado, aumentano la probabilità che la prole erediti due copie dello stesso allele raro. Tale incremento del rischio è spiegato dall’aumentata probabilità di identità per discendenza degli alleli. In termini formali, la prole di cugini primi presenta un coefficiente di inincrocio \(F = 1/16\), che quantifica la frazione attesa del genoma omozigote per identità per discendenza; per varianti deleterie rare, ciò si traduce in un aumento della frequenza di individui aa rispetto a popolazioni non consanguinee.
Mendel analizzò inizialmente incroci monoibridi per isolare il comportamento di un singolo carattere. Successivamente passò a incroci in cui due tratti venivano considerati contemporaneamente (incroci diibridi). Un esempio classico riguarda colore e forma del seme: il giallo domina sul verde e il liscio sul rugoso (Figura 07.13-03). Partendo da linee pure per entrambi i caratteri, una con fenotipi dominanti (YYRR) e una con fenotipi recessivi (yyrr), la generazione F1 risulta omogenea (YyRr) e manifesta i caratteri dominanti, indipendentemente dal fatto che gli alleli siano o meno associati. La chiave interpretativa emerge in F2, dopo autofecondazione delle piante F1. Se i due caratteri assortiscono indipendentemente, allora gli alleli per il colore segregano senza influenzare quelli per la forma. In termini meiotici, l’orientamento casuale dei bivalenti in metafase I è all’origine dell’indipendenza: ciascuna coppia di omologhi si dispone in modo indipendente dalle altre, generando combinazioni di alleli parentali nei gameti con probabilità uguali. Per un eterozigote doppio (YyRr), i tipi di gameti prodotti sono quattro e iso-frequenti: YR, Yr, yR, yr. La ricombinazione casuale di questi gameti genera in F2 quattro fenotipi: giallo-liscio, giallo-rugoso, verde-liscio e verde-rugoso. Il rapporto fenotipico atteso è 9:3:3:1, che si può derivare dal prodotto di due rapporti 3:1 indipendenti. In notazione probabilistica, indicando con A il fenotipo giallo e con B il fenotipo liscio, vale:
\[ \begin{aligned} P(A \cap B) &= \frac{3}{4} \cdot \frac{3}{4} = \frac{9}{16},\\ P(A \cap \neg B) &= \frac{3}{4} \cdot \frac{1}{4} = \frac{3}{16},\\ P(\neg A \cap B) &= \frac{1}{4} \cdot \frac{3}{4} = \frac{3}{16},\\ P(\neg A \cap \neg B) &= \frac{1}{4} \cdot \frac{1}{4} = \frac{1}{16}. \end{aligned} \]
Questo risultato è coerente con la seconda legge di Mendel, o legge dell’assortimento indipendente: ogni coppia di alleli si distribuisce nei gameti indipendentemente dalle altre coppie, purché i geni considerati si trovino su cromosomi differenti o, se sullo stesso, siano sufficientemente distanti da ricombinare frequentemente. La base cromosomica del principio è ancora una volta la metafase I: con n coppie di omologhi, il numero di assetti cromosomici distinti generabili per semplice orientamento indipendente è \(2^n\) (trascurando il crossing-over), il che contribuisce in modo sostanziale alla variabilità genetica. Nell’uomo, con 23 coppie di cromosomi, ciò equivale ad almeno \(2^{23}\) combinazioni possibili per la sola segregazione indipendente:
- incrocio diibrido tra linee pure (YYRR × yyrr) produce F1 uniformi (YyRr) con fenotipi dominanti;
- in F2, con assortimento indipendente, si osservano quattro fenotipi in rapporto 9:3:3:1, come previsto dalla moltiplicazione di due schemi 3:1;
- base meiotica: orientamento indipendente dei bivalenti in metafase I e, quando presente, ricombinazione per crossing-over che riduce l’associazione tra alleli sullo stesso cromosoma.
Image Gallery
Finché si rimane al livello concettuale, alleli e geni possono apparire entità astratte. È oggi assodato, tuttavia, che i fattori ipotizzati da Mendel coincidono con i geni localizzati sui cromosomi. Questi ultimi si separano durante la gametogenesi e si ricombinano alla fecondazione, fornendo il supporto fisico alle regole mendeliane. Nella meiosi I, i cromosomi omologhi di origine materna e paterna si appaiano, formano bivalenti e poi si disgiungono, distribuendo alleli alternativi in gameti distinti. Poiché gli omologhi spesso recano varianti alleliche differenti per molti loci, la segregazione meiotica determina la separazione ordinata di tali varianti. Si consideri, per esempio, una pianta di pisello eterozigote per il colore giallo del seme, Yy. Alla prima divisione meiotica, i cromosomi che portano rispettivamente l’allele Y e l’allele y vengono indirizzati verso poli opposti. Ne derivano gameti aploidi di due tipi, contenenti Y oppure y, ciascuno con probabilità \(0{,}5\). Nella successiva autofecondazione, la combinazione casuale dei gameti ristabilisce lo stato diploide e produce genotipi YY, Yy e yy con proporzioni attese coerenti con la legge della segregazione. La seconda legge di Mendel, relativa all’assortimento indipendente, trova spiegazione nella fisica dell’allineamento metafasico. Ogni bivalente si collega al fuso in modo autonomo e l’orientamento su piastra metafasica è indipendente per coppie diverse. Di conseguenza, ciascun gamete eredita una combinazione casuale di cromosomi materni e paterni e, pertanto, gli alleli di geni situati su cromosomi differenti segregano indipendentemente. Per una pianta YyRr, gli omologhi che portano gli alleli del colore dei semi e quelli della forma si orientano rispetto ai poli in modo stocastico; se il bivalente del colore presenta l’omologo con Y rivolto a un polo, nulla obbliga il bivalente della forma a condividere lo stesso orientamento. L’esito è una produzione equiprobabile di gameti YR, Yr, yR e yr, determinata dal caso dell’aggancio dei bivalenti ai microtubuli del fuso.
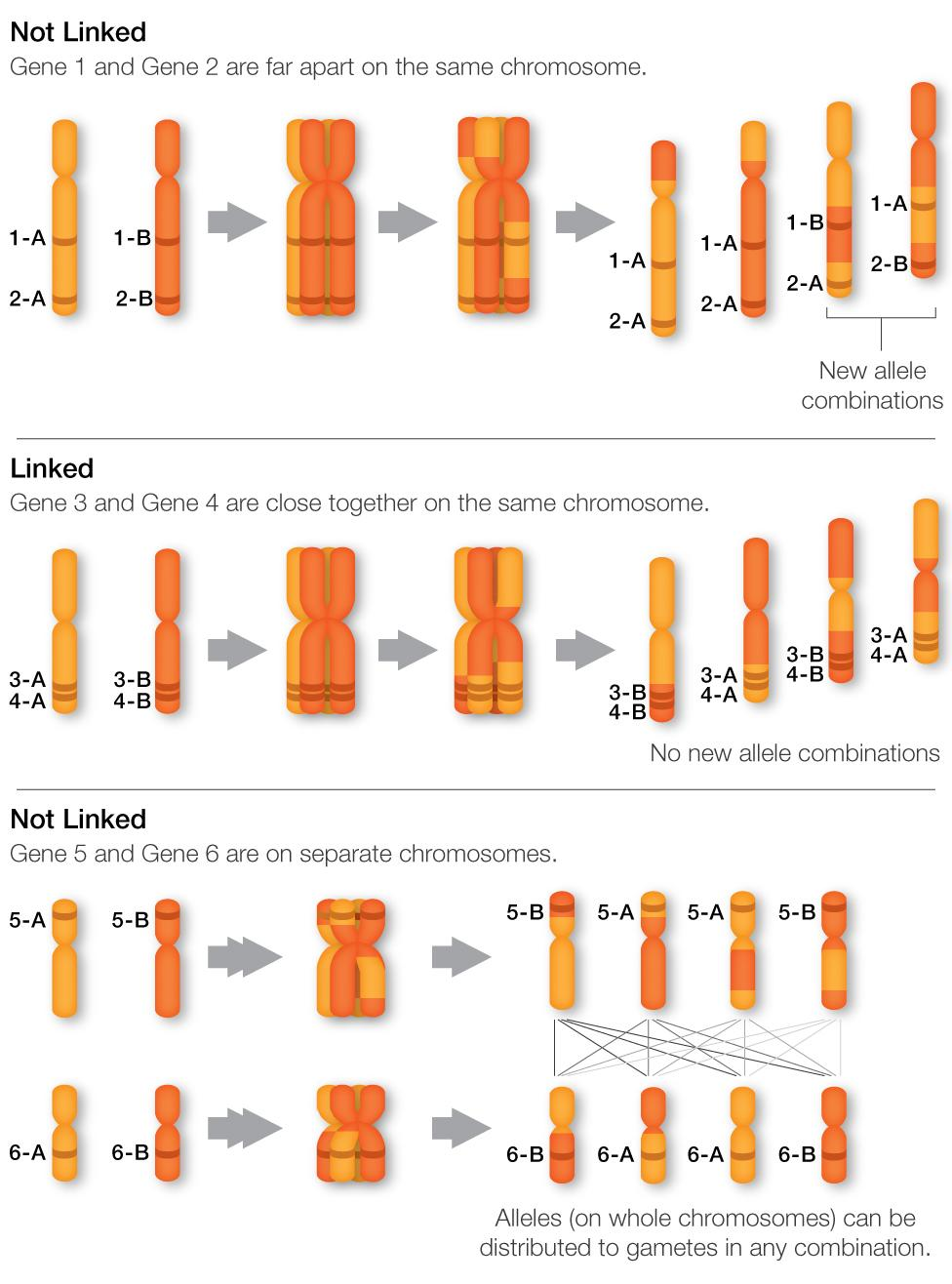
Mendel analizzò sette caratteri, ciascuno governato da un gene distinto. Sappiamo ora che molti di quei geni risiedono su cromosomi diversi, facilitando l’assortimento casuale osservato. Tuttavia, l’indipendenza di segregazione non richiede rigorosamente la collocazione su cromosomi differenti: due geni posti sullo stesso cromosoma, se separati da una distanza sufficiente, possono comunque assortire in modo indipendente grazie alla ricombinazione omologa (crossing over) che interviene in profase I. Quando i cromosomi omologhi duplicati si appaiano a formare bivalenti, tra cromatidi non fratelli si instaurano chiasmi, sedi di scambio di tratti di DNA. Tali scambi possono separare alleli inizialmente sullo stesso cromosoma, assegnandoli a gameti diversi (Figura 07.13-07). Un caso emblematico, oggi ben documentato, riguarda i geni per la forma del seme e per il colore del baccello nel pisello: benché colocalizzati sul medesimo cromosoma, sono così distanti da comportarsi come indipendenti nelle proporzioni fenotipiche, proprio per l’elevata probabilità di ricombinazione che li separa in meiosi. Non sempre, però, i geni si ereditano come prevede la seconda legge. Loci vicini sullo stesso cromosoma tendono a essere trasmessi insieme, fenomeno definito associazione genica (linkage). Negli umani, per esempio, l’allele responsabile della cecità rosso‑verde e quello dell’emofilia, entrambi in regioni adiacenti del cromosoma X, mostrano spesso coeredità. La frequenza di ricombinazione tra due loci, misurata come proporzione di gameti ricombinanti, fornisce una stima della loro distanza genetica: 1 centimorgan corrisponde a una probabilità di ricombinazione dell’1%. Questo principio ha consentito di costruire mappe genetiche in numerosi organismi, strumento che si è rivelato essenziale per identificare e isolare geni responsabili di patologie, come nel caso della mappatura e clonazione del gene della fibrosi cistica:
- distanza elevata sullo stesso cromosoma → alta probabilità di crossing over e comportamento quasi indipendente;
- distanza ridotta → bassa probabilità di scambio e coeredità dei caratteri;
- l’analisi delle frequenze ricombinanti consente l’ordinamento dei loci lungo il cromosoma.
Image Gallery
Le mutazioni sono alterazioni ereditabili della sequenza del DNA generate da errori di replicazione, agenti mutageni o processi endogeni. Dal punto di vista funzionale, esse si classificano in base all’impatto sull’attività genica. Le mutazioni con perdita di funzione riducono o aboliscono l’azione del gene o del suo prodotto (Figura 07.13-08). Un individuo omozigote per un allele con perdita di funzione manifesta spesso un fenotipo deviante rispetto al tipo più comune, sebbene in alcuni casi le differenze siano sottili. L’eterozigote che conserva un allele di tipo selvatico, nella maggior parte dei casi, produce quantitativi sufficienti di prodotto attivo per sostenere una funzione normale; per questo motivo molte mutazioni con perdita di funzione risultano recessive. Il classico esempio dei semi rugosi nel pisello illustra il meccanismo molecolare: il gene che controlla la forma del seme codifica un enzima coinvolto nella conversione degli zuccheri in amido ramificato. L’allele dominante R consente la sintesi dell’enzima attivo, mentre l’allele recessivo r produce un enzima non funzionale. Le piante rr accumulano zuccheri solubili e sintetizzano meno amido; durante l’essiccamento questo sbilanciamento osmotico induce il caratteristico aspetto rugoso. Non sorprende che molte varietà commerciali di piselli dolci derivino da alleli con perdita di funzione simili a quelli analizzati negli esperimenti classici. Non tutte le perdite di funzione sono recessive. In presenza di aploinsufficienza, metà della dose normale non basta a garantire la funzione e la mutazione si manifesta in eterozigosi. Altre volte, una proteina mutata può interferire con quella normale esercitando un effetto dominante negativo, come osservato per proteine strutturali oligomeriche. Vi è poi la classe opposta: le mutazioni con guadagno di funzione, che aumentano l’attività del gene o promuovono espressione e/o attività in contesti inappropriati; queste, in genere, sono dominanti (Figura 07.13-08). Un esempio rilevante riguarda mutazioni attivanti di Ras, che bloccano la proteina in uno stato GTP‑legato costitutivamente attivo. Poiché Ras integra segnali proliferativi, la sua attivazione inappropriata promuove divisione cellulare anche in assenza di stimoli esterni, contribuendo alla trasformazione neoplastica; mutazioni di questo tipo sono riscontrate in circa il 30% dei tumori umani.
Image Gallery
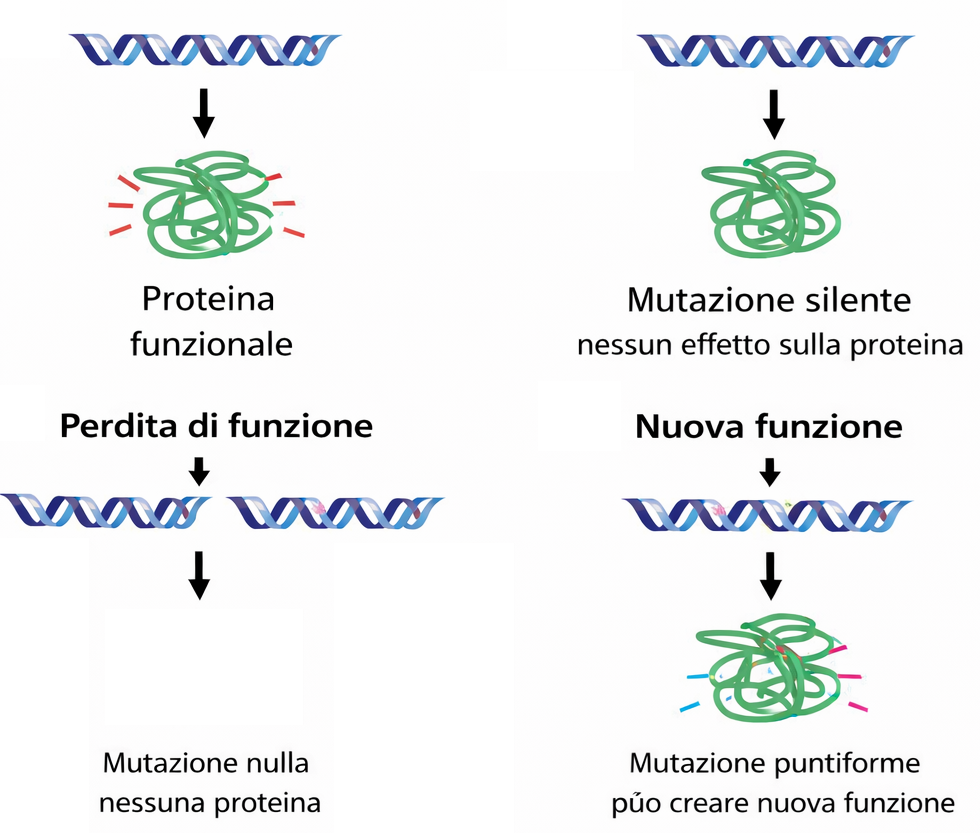
Image Gallery
La variabilità genetica originata dalle mutazioni costituisce il materiale su cui agisce la selezione naturale. Mutazioni che conferiscono un vantaggio tendenzialmente aumentano di frequenza, mentre quelle che compromettono sopravvivenza o fertilità tendono a scomparire. La gran parte delle nuove varianti è neutra o sfavorevole. Se una mutazione deleteria è dominante, di solito viene rapidamente rimossa perché il fenotipo negativo compare già in eterozigosi e riduce la probabilità di trasmissione alla progenie. Diversa è la dinamica per le mutazioni deleterie recessive: l’eterozigote è fenotipicamente sano, per cui l’allele può persistere “nascosto” alla selezione e propagarsi nella popolazione. In una popolazione reale, l’equilibrio tra l’introduzione di nuove mutazioni e la loro rimozione attraverso la selezione contro gli omozigoti deleteri conduce a una frequenza stazionaria approssimabile, per un allele recessivo con effetto selettivo \(s\) sugli omozigoti e tasso di mutazione \(\mu\), da: \[ q_{\mathrm{eq}} \approx \sqrt{\frac{\mu}{s}}. \] Tale relazione, pur nella sua semplicità, spiega perché alleli recessivi patogenici possano mantenersi a frequenze non trascurabili anche quando gli omozigoti presentano una ridotta fitness. Fattori demografici e sociali, come l’isolamento riproduttivo o la consanguineità, possono aumentare la probabilità che due portatori condividano lo stesso allele recessivo e abbiano figli affetti. Un esempio didattico: una forma comune di ipoacusia ereditaria infantile dovuta a mutazioni in una proteina di giunzione comunicante si manifesta in circa 1 su 2500 nati, mentre la frequenza di portatori eterozigoti può aggirarsi intorno a 1 su 40. Sebbene gli individui clinicamente affetti siano rari, la presenza di portatori è relativamente elevata proprio perché l’allele deleterio rimane occultato in eterozigosi:
- le mutazioni recessive permangono perché non sono “viste” dalla selezione in eterozigosi;
- il bilancio mutazione‑selezione stabilisce la frequenza a lungo termine degli alleli deleteri;
- la struttura della popolazione (dimensione efficace, migrazione, assortimento) modula le probabilità di incontro tra portatori.


