La struttura dei cromosomi negli organismi eucarioti
La quantità di informazione necessaria a specificare anche un microrganismo semplice è enorme e richiede lunghe molecole di DNA. Nell’essere umano, ogni cellula somatica contiene circa 2 metri di DNA, compattati all’interno di un nucleo di appena 5–8 μm di diametro. In termini di rapporto lineare, ciò equivale a ridurre una corda lunga chilometri a un volume subcellulare, con un fattore di compattazione dell’ordine di \(10^{5}\)–\(10^{6}\). Questa compressione estrema è ottenuta senza compromettere l’accesso selettivo al DNA da parte degli apparati che devono leggerlo, duplicarlo e ripararlo.
Nelle cellule eucariotiche, i lunghi filamenti di DNA a doppia elica sono organizzati in cromosomi. L’architettura cromosomica non si limita a “far entrare” il DNA nel nucleo: deve consentirne la duplicazione accurata, l’assemblaggio e la segregazione controllata nelle cellule figlie durante la divisione. Questa impresa è resa possibile dalla cromatina, un complesso dinamico di DNA e proteine che ripiega la molecola in anelli, spire e domini topologici. Le proteine che mediano tale organizzazione includono gli istoni, che formano i nucleosomi, proteine non istoniche regolatrici, complessi SMC come coesine e condensine che governano l’architettura su larga scala, e topoisomerasi che modulano la superavvolgimento. Nonostante l’elevato grado di compattazione, la cromatina rimane funzionalmente accessibile, permettendo la regolazione fine dell’espressione genica, la replicazione e i processi di riparo.
Nei procarioti, in genere, i geni sono raccolti su un’unica molecola di DNA circolare, organizzata nel nucleoide. Anche qui intervengono proteine che condensano il DNA, come HU, IHF, Fis e H-NS, e l’assetto è regolato da superavvolgimento e ponti proteici; tuttavia, la struttura risultante differisce profondamente da quella dei cromosomi eucariotici. Poiché i meccanismi di condensazione procarioti non sono sovrapponibili a quelli eucarioti, l’analisi che segue si concentra sulla struttura e funzione dei cromosomi degli eucarioti.
Nei eucarioti il DNA nucleare è ripartito in più cromosomi
Negli eucarioti, compresa la specie umana, il DNA nucleare è distribuito in cromosomi distinti. Nell’uomo si riconoscono 23 o 24 tipi cromosomici, a seconda del sesso: nella femmina 23 categorie (22 autosomi più il cromosoma X), nel maschio 24 (gli stessi 22 autosomi, più X e Y). Ciascun cromosoma corrisponde a una singola molecola di DNA lineare di grande lunghezza, associata a un insieme di proteine che ne orchestrano il ripiegamento in una struttura compatta e ordinata. Il complesso DNA–proteine è definito cromatina. Oltre alle proteine deputate all’impacchettamento, su ogni cromosoma si assemblano fattori coinvolti nell’espressione genica, nella replicazione, nel controllo della qualità del DNA e nella sua riparazione.
Le cellule somatiche umane possiedono due copie di ciascun cromosoma, una di origine materna e una di origine paterna, formando coppie di omologhi. Fanno eccezione i gameti, che sono aploidi \((n=23)\), e cellule altamente specializzate prive di nucleo, come gli eritrociti maturi. La coppia dei cromosomi sessuali è l’unica non costituita da omologhi identici nel maschio, che presenta un X e un Y; nella femmina sono presenti due cromosomi X. L’insieme di tutte le sequenze nucleari contenute in una singola serie cromosomica costituisce il genoma aploide; nell’uomo ammonta a circa \(3{,}2 \times 10^{9}\) coppie di nucleotidi. Oltre al DNA nucleare, le cellule umane contengono anche il DNA mitocondriale, circolare e in molte copie, ereditato in prevalenza per via materna.
L’identificazione dei singoli cromosomi si avvale di tecniche citogenetiche complementari. La ibridazione in situ fluorescente con sonde specifiche “vernicia” ciascun cromosoma con combinazioni di fluorocromi (Figura 02.03-01). I metodi di bandeggio tradizionali, come il bandeggio G o R, sfruttano coloranti che evidenziano regioni con differente composizione in basi (ricche in A–T o in G–C), producendo un pattern regolare di bande riproducibile che consente di distinguere e numerare i cromosomi. L’ordinamento dei 46 cromosomi in una tavola convenzionale costituisce il cariotipo umano (Figura 02.03-01). Variazioni strutturali, quali delezioni, duplicazioni, traslocazioni o inversioni, alterano i pattern attesi e possono essere riconosciute con questi approcci. L’analisi cariotipica è uno strumento chiave per rilevare anomalie associate a patologie ereditarie e a neoplasie (Figura 02.03-02).
Image Gallery

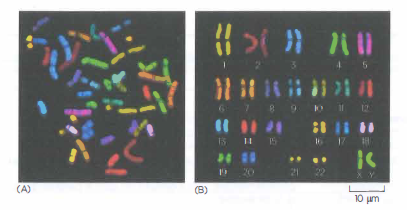
Cromosomi umani multicolori e cariotipo
È possibile “colorare” ogni cromosoma umano di un colore diverso per riuscire a identificarlo al microscopio ottico senza ambiguità. Questi cromosomi sono stati isolati da una cellula con il nucleo in divisione (in mitosi) e quindi hanno una struttura estremamente compatta (condensata). I cromosomi multicolori si ottengono trattandoli con una serie di molecole di DNA umano marcate con una combinazione di coloranti fluorescenti. Per esempio, le molecole di DNA derivate dal cromosoma 1 si marcano con una certa combinazione di coloranti, le molecole di DNA derivate dal cromosoma 2 con un’altra e così via. Dato che il DNA marcato va a ibridarsi, cioè a formare accoppiamenti di basi, solo con il cromosoma di origine, ogni cromosoma si marca diversamente. In questi esperimenti i cromosomi subiscono trattamenti che, pur mantenendo la struttura relativamente intatta, separano il DNA a doppia elica nei due filamenti per far appaiare ciascuno di essi con il DNA marcato a singolo filamento. (A) Cromosomi visualizzati così come fuoriescono dalla cellula lisata.(B) Gli stessi cromosomi allineati artificialmente in ordine numerico. Questa disposizione della serie completa di cromosomi si chiama cariotipo.
Immagine tratta liberamente da Internet. Se viola i tuoi diritti, contattaci.
Image Gallery
Cromosomi umani multicolori e cariotipo
È possibile “colorare” ogni cromosoma umano di un colore diverso per riuscire a identificarlo al microscopio ottico senza ambiguità. Questi cromosomi sono stati isolati da una cellula con il nucleo in divisione (in mitosi) e quindi hanno una struttura estremamente compatta (condensata). I cromosomi multicolori si ottengono trattandoli con una serie di molecole di DNA umano marcate con una combinazione di coloranti fluorescenti. Per esempio, le molecole di DNA derivate dal cromosoma 1 si marcano con una certa combinazione di coloranti, le molecole di DNA derivate dal cromosoma 2 con un’altra e così via. Dato che il DNA marcato va a ibridarsi, cioè a formare accoppiamenti di basi, solo con il cromosoma di origine, ogni cromosoma si marca diversamente. In questi esperimenti i cromosomi subiscono trattamenti che, pur mantenendo la struttura relativamente intatta, separano il DNA a doppia elica nei due filamenti per far appaiare ciascuno di essi con il DNA marcato a singolo filamento. (A) Cromosomi visualizzati così come fuoriescono dalla cellula lisata.(B) Gli stessi cromosomi allineati artificialmente in ordine numerico. Questa disposizione della serie completa di cromosomi si chiama cariotipo.
Immagine tratta liberamente da Internet. Se viola i tuoi diritti, contattaci.
Image Gallery
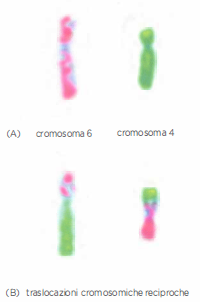
Cromosomi anomali e traslocazioni
Cromosomi anomali sono associati ad alcuni difetti genetici ereditari. (A) Due cromosomi umani normali, il cromosoma 6 e il cromosoma 4, sono stati marcati come descritto nella Figura 02.03-01. (B) In un individuo con una traslocazione cromosomica reciproca, un segmento di un cromosoma è stato scambiato con un segmento di un altro. Tali traslocazioni cromosomiche sono un evento frequente nelle cellule tumorali.
Immagine tratta liberamente da Internet. Se viola i tuoi diritti, contattaci.
I cromosomi come contenitori e organizzatori dell’informazione genetica
Il ruolo cardine dei cromosomi è quello di organizzare e trasmettere i geni, unità funzionali dell’ereditarietà. Operativamente, un gene è un segmento di DNA che specifica un prodotto funzionale, generalmente una proteina oppure una molecola di RNA. Molti trascritti di RNA vengono tradotti in proteine; altri, invece, agiscono come RNA non codificanti con funzioni strutturali, catalitiche o regolatorie (Figura 02.03-17), contribuendo al controllo dell’espressione genica e all’architettura della cromatina.
Il complesso delle informazioni contenute in tutti i cromosomi di una cellula o di un organismo è il genoma. Sono oggi disponibili le sequenze complete di migliaia di specie, da Escherichia coli all’uomo. Benché organismi più complessi tendano a possedere più geni, la relazione non è lineare. Alcuni batteri a genoma ridotto contano poche centinaia di geni, mentre il genoma umano comprende dell’ordine di decine di migliaia di geni codificanti e numerosi RNA non codificanti. Genomi particolarmente compatti, come quelli batterici o di eucarioti unicellulari quali Saccharomyces cerevisiae, presentano cromosomi con alta densità genica e spazi intergenici ridotti (Figura 02.03-03). In contrasto, i cromosomi di molti eucarioti superiori, uomo incluso, contengono ampie regioni intergeniche e introniche, oltre a sequenze necessarie per la regolazione, la replicazione e la stabilità del DNA (Figura 02.03-04).
Il cosiddetto “DNA non codificante” non equivale a materiale privo di funzione. Gran parte di queste sequenze include elementi regolatori (promotori, enhancer, insulator), RNA non codificanti, DNA ripetuto e residui di elementi trasponibili. Confronti genomici tra specie imparentate mostrano che una frazione rilevante di questo DNA è conservata, suggerendo ruoli biologici importanti, talvolta non ancora pienamente chiariti. La variabilità della quantità di DNA non codificante contribuisce alla cosiddetta “paradosso del C‑value”, per cui la dimensione del genoma non correla in modo semplice con la complessità dell’organismo.
In generale, all’aumentare della complessità biologica cresce la dimensione genomica, ma esistono ampie eccezioni. Il genoma umano è circa 200 volte più grande di quello di S. cerevisiae, ma risulta molto più piccolo di quello di talune piante o di alcune amebe. Anche il numero di cromosomi varia notevolmente tra specie: l’uomo possiede 46 cromosomi (in cellule diploidi), una specie di cervide ne presenta solo 7, mentre in alcune carpe si superano le 100 unità. Persino organismi strettamente imparentati e con dimensioni genomiche simili possono differire sostanzialmente per numero e lunghezza dei cromosomi (Figura 02.03-05). Queste discrepanze riflettono una storia evolutiva fatta di duplicazioni, fusioni e fissioni cromosomiche, mobilità di elementi genetici e selezione naturale, che hanno rimodellato nel tempo genomi e cariotipi:
- Fattori che contribuiscono alle dimensioni genomiche: introni e spazi intergenici estesi;
- espansioni di elementi ripetitivi ed elementi trasponibili;
- duplicazioni segmentali e intere poliploidizzazioni;
- diversi regimi selettivi e tassi di mutazione che influenzano la ritenzione o la perdita di sequenze;
- riorganizzazioni cromosomiche che alterano numero e struttura dei cromosomi.
In sintesi, i cromosomi non sono semplici contenitori, ma piattaforme organizzative che consentono di integrare compattazione, accessibilità e controllo dell’informazione genetica in modo compatibile con le esigenze della vita cellulare.
Image Gallery
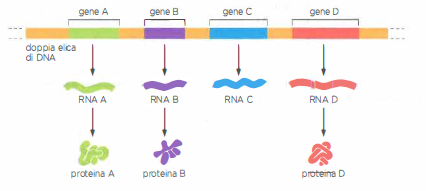
Geni ed espressione genica
La maggior parte dei geni contiene informazioni per produrre le proteine. Come esposto, ogni gene che codifica proteine viene impiegato nella sintesi di molecole di RNA, che a loro volta pilotano la sintesi di molecole proteiche specifiche. Si noti che per una certa quantità di geni il prodotto finale è la stessa molecola di RNA, come mostrato qui per il gene C. In questi casi l’espressione genica è completa una volta che la sequenza nucleotidica del DNA è stata trascritta nella sequenza nucleotidica del suo RNA.
Immagine tratta liberamente da Internet. Se viola i tuoi diritti, contattaci.
Image Gallery
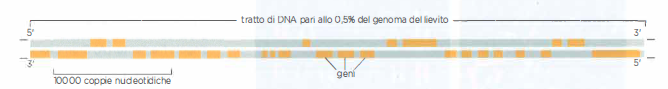
Geni e cromosomi nel lievito
Nel lievito, i geni sono strettamente impacchettati lungo i cromosomi. Questa figura mostra una piccola regione della doppia elica del DNA in un cromosoma del lievito S. cerevisiae. Il genoma di S. cerevisiae contiene circa 12,5 milioni di coppie nucleotidiche e 6600 geni, distribuiti su 16 cromosomi. Si noti che, per ciascun gene, solo uno dei due filamenti di DNA codifica effettivamente le informazioni per produrre una molecola di RNA. Questa regione codificante può essere su entrambi i filamenti, come indicato dalle barre arancioni. Tuttavia, si ritiene che ogni “gene” includa sia il “filamento codificante” sia il suo complementare. L’alta densità dei geni è caratteristica di S. cerevisiae.
Immagine tratta liberamente da Internet. Se viola i tuoi diritti, contattaci.
Image Gallery

Gene β-globina
In molti eucarioti, i geni includono un eccesso di DNA inframezzato, non codificante. Qui è rappresentata la sequenza nucleotidica del gene della β-globina umana. Questo gene trasporta l’informazione che specifica la sequenza amminoacidica di uno dei due tipi di subunità presenti nell’emoglobina, una proteina che trasporta l’ossigeno nel sangue. È mostrata solo la sequenza del filamento codificante: il filamento non codificante ospita la sequenza complementare. A partire dall’estremità 5′, tale sequenza viene letta da sinistra a destra, come qualsiasi libro di testo. I segmenti della sequenza del DNA che codificano la sequenza amminoacidica della β-globina sono evidenziati in giallo. Vedremo successivamente come queste informazioni vengano trascritte e tradotte per produrre una proteina β-globina completa.
Immagine tratta liberamente da Internet. Se viola i tuoi diritti, contattaci.
Image Gallery

Numero cromosomi specie affini
Due specie strettamente imparentate possono essere simili per quanto riguarda le dimensioni del genoma, ma avere un numero di cromosomi molto diverso. Nell’evoluzione del muntjac indiano, i cromosomi che inizialmente erano separati, e che sono rimasti separati nella specie cinese, si sono fusi senza produrre un effetto significativo sull’animale.
Immagine tratta liberamente da Internet. Se viola i tuoi diritti, contattaci.
Sequenze funzionali del DNA per duplicazione e corretta segregazione cromosomica
Affinché un cromosoma sia operativo, non è sufficiente che il DNA contenga i geni: la molecola deve potersi copiare con accuratezza e le copie devono essere ripartite in modo equo tra le cellule figlie a ogni divisione. Tali eventi si svolgono secondo un programma temporale coordinato, il ciclo cellulare, schematicamente illustrato nella (Figura 02.03-06). In questa sede si considerano due fasi cardine: l’interfase, in cui avviene la duplicazione dei cromosomi, e la mitosi, durante la quale i cromosomi replicati vengono separati nei due nuclei figli.
Nell’interfase, la cromatina si presenta distesa e forma una rete filamentosa poco distinguibile al microscopio ottico (Figura 02.03-18); in tale configurazione, i cromosomi sono detti interfasici. È in questo intervallo che si realizza la replicazione del DNA. Sequenze specializzate assicurano che la duplicazione proceda in modo efficiente. In particolare, le origini di replicazione sono siti di avvio della sintesi del DNA: i cromosomi eucariotici ne possiedono numerose, consentendo la replicazione simultanea di più segmenti e accelerando l’intero processo (Figura 02.03-07). Alle estremità dei cromosomi si trovano i telomeri, costituiti da brevi ripetizioni nucleotidiche tandem e da proteine specifiche che proteggono le fini cromosomiche dalla fusione e dal riconoscimento come rotture del DNA, oltre a facilitare la copia completa delle estremità attraverso l’azione della telomerasi.
Un terzo elemento chiave è il centromero, una regione di DNA e cromatina specializzata essenziale per la suddivisione dei cromosomi duplicati durante la fase M (Figura 02.03-07). Con l’ingresso in mitosi, la cromatina si condensa progressivamente fino a generare i cromosomi mitotici altamente compatti (Figura 02.03-08), facilmente osservabili al microscopio (Figura 02.03-18). Il centromero nuclea la formazione del cinetocore, complesso proteico che si collega ai microtubuli del fuso e coordina l’attrazione dei cromatidi fratelli verso poli opposti, garantendo che ciascuna cellula figlia riceva una copia completa del patrimonio cromosomico (Figura 02.03-06):
- Origini di replicazione: definiscono i punti d’avvio della duplicazione e distribuite in più copie per cromosoma, riducono i tempi complessivi della sintesi del DNA;
- Telomeri: ripetizioni terminali e complessi proteici associati che risolvono il problema della replicazione delle estremità e proteggono i capi cromosomici da degradazione e riarrangiamenti;
- Centromeri: domini cromatinici specializzati, sede del cinetocore, indispensabili per l’aggancio al fuso e la segregazione fedele dei cromatidi fratelli.
Image Gallery
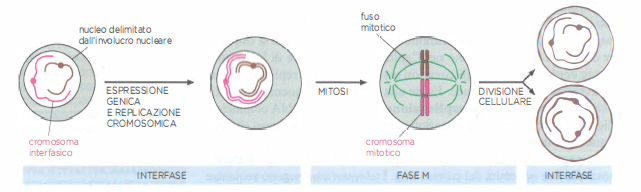
Ciclo cellulare
Nelle cellule che proliferano la replicazione e la divisione dei cromosomi avvengono secondo una serie di stadi prevedibili, detti nell’insieme ciclo cellulare. Durante l’interfase, la cellula esprime molti dei suoi geni e, durante parte di questa fase, duplica i cromosomi. Una volta che il DNA ha finito di replicarsi, la cellula può entrare nella fase M, in cui avviene la divisione cellulare o mitosi. In questo stadio i cromosomi si condensano, l’espressione genica è in gran parte sospesa, l’involucro nucleare si frammenta e si forma il fuso mitotico, composto da microtubuli e altre proteine. Il fuso mitotico cattura i cromosomi condensati e ne trascina una serie completa a ciascun polo della cellula; intorno a ogni corredo cromosomico si riforma la membrana nucleare e, nell’ultima tappa della fase M, la cellula si divide in due cellule figlie. Per semplicità nello schema sono indicati solo due cromosomi diversi.
Immagine tratta liberamente da Internet. Se viola i tuoi diritti, contattaci.
Image Gallery
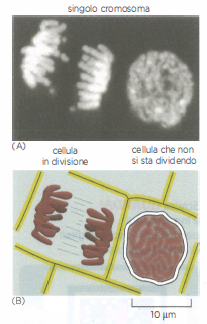
Cromosomi e divisione cellulare
I cromosomi si rendono visibili quando la cellula eucariote si prepara a dividersi. (A) Due cellule vegetali adiacenti fotografate al microscopio ottico a fluorescenza. Il DNA è stato trattato con un colorante fluorescente (DAPI) che lo lega specificamente. Il DNA si trova nei cromosomi, che diventano visibili al microscopio ottico solo quando si addensano in preparazione alla divisione cellulare, come si vede a sinistra. Per chiarezza, un singolo cromosoma è stato ombreggiato (in marrone) nella cellula in divisione. La cellula a destra, che non si sta dividendo, contiene gli stessi cromosomi, che però non si possono identificare al microscopio ottico in questa fase del ciclo vitale perché il DNA ha una conformazione molto più distesa. (B) Profilo schematico delle due cellule e dei loro cromosomi.
Immagine tratta liberamente da Internet. Se viola i tuoi diritti, contattaci.
Image Gallery
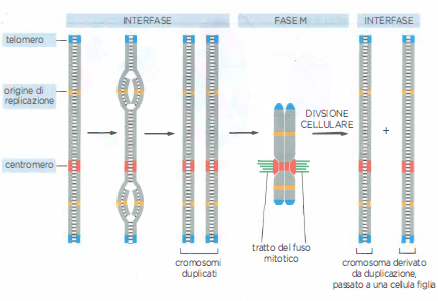
Elementi essenziali del cromosoma
Tre sono gli elementi che devono essere presenti nella sequenza del DNA perché il cromosoma eucariote possa replicarsi e poi segregare al momento della mitosi. Ogni cromosoma ha origini di replicazione multiple, un centromero e due telomeri. Nella figura è schematizzata la serie di eventi che interessano un cromosoma tipico durante il ciclo cellulare. Durante l’interfase il DNA si replica a partire dalle origini di replicazione in entrambe le direzioni e per tutta la sua lunghezza. Nella fase M il centromero attacca ognuno dei cromosomi duplicati al fuso mitotico, in modo che, quando la cellula si divide, una copia venga distribuita a ognuna delle cellule figlie. Prima della divisione cellulare, il centromero serve anche a mantenere insieme i cromosomi duplicati finché non sono pronti a separarsi. I telomeri contengono sequenze di DNA che consentono la duplicazione completa delle estremità cromosomiche.
Immagine tratta liberamente da Internet. Se viola i tuoi diritti, contattaci.
Image Gallery
Elementi essenziali del cromosoma
Tre sono gli elementi che devono essere presenti nella sequenza del DNA perché il cromosoma eucariote possa replicarsi e poi segregare al momento della mitosi. Ogni cromosoma ha origini di replicazione multiple, un centromero e due telomeri. Nella figura è schematizzata la serie di eventi che interessano un cromosoma tipico durante il ciclo cellulare. Durante l’interfase il DNA si replica a partire dalle origini di replicazione in entrambe le direzioni e per tutta la sua lunghezza. Nella fase M il centromero attacca ognuno dei cromosomi duplicati al fuso mitotico, in modo che, quando la cellula si divide, una copia venga distribuita a ognuna delle cellule figlie. Prima della divisione cellulare, il centromero serve anche a mantenere insieme i cromosomi duplicati finché non sono pronti a separarsi. I telomeri contengono sequenze di DNA che consentono la duplicazione completa delle estremità cromosomiche.
Immagine tratta liberamente da Internet. Se viola i tuoi diritti, contattaci.
Image Gallery
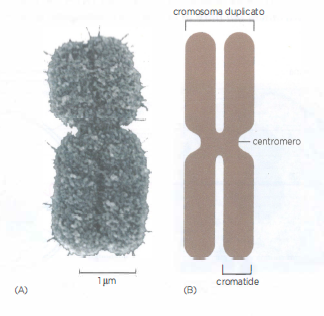
Cromosoma mitotico duplicato
Un tipico cromosoma mitotico duplicato è ipercompatto. Poiché il DNA viene replicato durante l’interfase, ogni cromosoma mitotico duplicato contiene due molecole di DNA figlie identiche (Figura 02.03-07). Ciascuna di queste molecole lunghissime di DNA, insieme alle proteine associate, si chiama cromatide. Una volta separati, i due cromatidi fratelli vengono considerati cromosomi singoli. (A) Un cromosoma mitotico fotografato al microscopio elettronico a scansione. I due cromatidi sono accorpati: la zona di strozzatura indica la posizione del centromero. (B) Rappresentazione schematica di un cromosoma mitotico.
Immagine tratta liberamente da Internet. Se viola i tuoi diritti, contattaci.
Image Gallery
Cromosomi e divisione cellulare
I cromosomi si rendono visibili quando la cellula eucariote si prepara a dividersi. (A) Due cellule vegetali adiacenti fotografate al microscopio ottico a fluorescenza. Il DNA è stato trattato con un colorante fluorescente (DAPI) che lo lega specificamente. Il DNA si trova nei cromosomi, che diventano visibili al microscopio ottico solo quando si addensano in preparazione alla divisione cellulare, come si vede a sinistra. Per chiarezza, un singolo cromosoma è stato ombreggiato (in marrone) nella cellula in divisione. La cellula a destra, che non si sta dividendo, contiene gli stessi cromosomi, che però non si possono identificare al microscopio ottico in questa fase del ciclo vitale perché il DNA ha una conformazione molto più distesa. (B) Profilo schematico delle due cellule e dei loro cromosomi.
Immagine tratta liberamente da Internet. Se viola i tuoi diritti, contattaci.
Image Gallery
Ciclo cellulare
Nelle cellule che proliferano la replicazione e la divisione dei cromosomi avvengono secondo una serie di stadi prevedibili, detti nell’insieme ciclo cellulare. Durante l’interfase, la cellula esprime molti dei suoi geni e, durante parte di questa fase, duplica i cromosomi. Una volta che il DNA ha finito di replicarsi, la cellula può entrare nella fase M, in cui avviene la divisione cellulare o mitosi. In questo stadio i cromosomi si condensano, l’espressione genica è in gran parte sospesa, l’involucro nucleare si frammenta e si forma il fuso mitotico, composto da microtubuli e altre proteine. Il fuso mitotico cattura i cromosomi condensati e ne trascina una serie completa a ciascun polo della cellula; intorno a ogni corredo cromosomico si riforma la membrana nucleare e, nell’ultima tappa della fase M, la cellula si divide in due cellule figlie. Per semplicità nello schema sono indicati solo due cromosomi diversi.
Immagine tratta liberamente da Internet. Se viola i tuoi diritti, contattaci.
Organizzazione spaziale dei cromosomi nell’interfase
I cromosomi interfasici, più lunghi e sottili dei loro corrispettivi mitotici, non sono disposti a caso nel nucleo, ma mostrano un’organizzazione gerarchica. Sebbene dinamici, essi occupano territori nucleari distinti che limitano l’intreccio eccessivo tra cromosomi diversi (Figura 02.03-09). Interazioni con la membrana nucleare interna e con la lamina nucleare, un reticolo di proteine fibrose, contribuiscono al posizionamento e al confinamento dei cromosomi all’interno dei rispettivi domini, favorendo al contempo regolazioni funzionali quali la repressione o l’attivazione di regioni geniche attraverso associazioni con domini della lamina.
Una manifestazione evidente di tale organizzazione è il nucleolo, ben visibile al microscopio ottico come corpo subnucleare (Figura 02.03-10). Durante l’interfase, segmenti provenienti da più cromosomi che contengono i geni per gli RNA ribosomiali si riuniscono a formare questa struttura. Nell’uomo, centinaia di copie dei geni rDNA sono raggruppate in 10 regioni organizzatrici nucleolari, collocate vicino alle estremità di cinque coppie di cromosomi acrocentrici (Figura 02.03-10). Nel nucleolo, l’RNA polimerasi I trascrive gli rRNA, che vengono poi processati e assemblati con proteine ribosomiali per formare le subunità dei ribosomi, macchine catalitiche deputate alla sintesi proteica. Gli rRNA apportano contributi sia strutturali sia catalitici all’interno del ribosoma, coordinando l’allineamento dei substrati e la formazione del legame peptidico.
Image Gallery
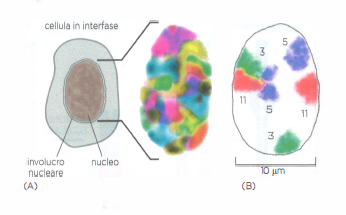
Cromosomi interfasici
Nel nucleo i cromosomi interfasici occupano specifiche regioni diverse. Per distinguere tramite colorazione specifica i singoli cromosomi interfasici nella cellula umana, si impiegano sonde (probes) a DNA accoppiate a coloranti fluorescenti diversi. (A) Con il microscopio a fluorescenza si vede che il nucleo è riempito con un mosaico di colori distinti. (B) Per evidenziare le loro posizioni, vengono individuate tre serie di cromosomi: cromosoma 3, 5 e 11. Si noti che le coppie di cromosomi omologhi, come le due copie del cromosoma 3, non si trovano generalmente nella stessa posizione.
Immagine tratta liberamente da Internet. Se viola i tuoi diritti, contattaci.
Image Gallery
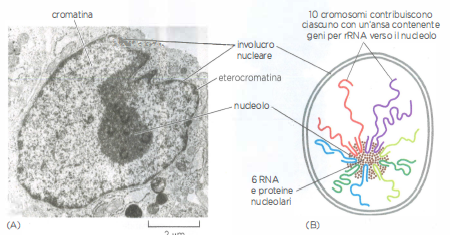
Nucleolo
Il nucleolo è la struttura più cospicua del nucleo interfascico. (A) Sezione ultrasottile di un nucleo di fibroblasto umano al microscopio elettronico. Il nucleo è circondato dall’involucro nucleare. Al suo interno, la cromatina si presenta come una massa diffusa e marmorizzata, con regioni più dense, dette eterocromatina (di colorazione scura). L’eterocromatina è povera di geni e si dispone principalmente alla periferia del nucleo, subito sotto l’involucro nucleare. L’ampia regione scura centrale è il nucleolo, che contiene i geni per l’RNA ribosomiale. (B) Illustrazione schematica che mostra come i geni per l’RNA ribosomiale, raggruppati vicino alle estremità di cinque diversi cromosomi umani (13, 14, 15, 21 e 22), si uniscono per formare il nucleolo, un sottocompartimento biochimico prodotto dall’aggregazione di una serie di macromolecole: DNA, RNA e proteine.
Immagine tratta liberamente da Internet. Se viola i tuoi diritti, contattaci.
Image Gallery
Nucleolo
Il nucleolo è la struttura più cospicua del nucleo interfascico. (A) Sezione ultrasottile di un nucleo di fibroblasto umano al microscopio elettronico. Il nucleo è circondato dall’involucro nucleare. Al suo interno, la cromatina si presenta come una massa diffusa e marmorizzata, con regioni più dense, dette eterocromatina (di colorazione scura). L’eterocromatina è povera di geni e si dispone principalmente alla periferia del nucleo, subito sotto l’involucro nucleare. L’ampia regione scura centrale è il nucleolo, che contiene i geni per l’RNA ribosomiale. (B) Illustrazione schematica che mostra come i geni per l’RNA ribosomiale, raggruppati vicino alle estremità di cinque diversi cromosomi umani (13, 14, 15, 21 e 22), si uniscono per formare il nucleolo, un sottocompartimento biochimico prodotto dall’aggregazione di una serie di macromolecole: DNA, RNA e proteine.
Immagine tratta liberamente da Internet. Se viola i tuoi diritti, contattaci.
Il DNA cromosomico mantiene un elevato grado di compattazione
Nelle cellule eucariotiche il DNA è fortemente impacchettato sia in interfase sia in mitosi. Un confronto numerico chiarisce l’ordine di grandezza del fenomeno: il cromosoma 1 umano contiene circa 249 milioni di coppie di basi, che corrisponderebbero a una lunghezza distesa di circa 8,5 cm; in metafase, la stessa unità cromosomica misura pochi micrometri, implicando una compressione complessiva dell’ordine di \(10^4\). Le proteine associate al DNA orchestrano una serie di ripiegamenti e avvolgimenti modulari, generando livelli successivi di compattazione. Durante l’interfase, la cromatina è in media circa 15–20 volte meno condensata rispetto allo stato mitotico (Figura 02.03-11), pur mantenendo una notevole compattazione rispetto al DNA nudo.
La struttura cromosomica è tuttavia intrinsecamente dinamica: deve consentire un accesso rapido e selettivo a sequenze specifiche per processi quali replicazione, riparazione ed espressione genica. Complessi proteici dedicati, inclusi rimodellatori della cromatina ATP-dipendenti e topoisomerasi, modulano localmente densità e topologia della fibra, assicurando un bilanciamento continuo tra compattazione e accessibilità.
Image Gallery
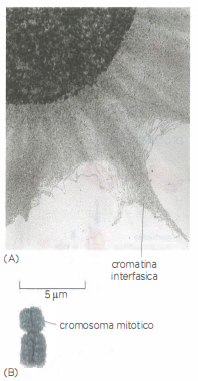
Cromosoma interfascico
Nel cromosoma interfascico il DNA è meno compatto che nel cromosoma mitotico. (A) Immagine al microscopio elettronico che mostra un enorme groviglio di cromatina (DNA e proteine associate) mentre sta uscendo da un nucleo interfascico lisato. (B) Rappresentazione schematica di un cromosoma mitotico umano della stessa scala.
Immagine tratta liberamente da Internet. Se viola i tuoi diritti, contattaci.
Il nucleosoma costituisce il modulo elementare della cromatina eucariotica
Le proteine che si associano al DNA nucleare appartengono a due grandi classi: gli istoni e una vasta collezione di proteine non istoniche. Nel loro insieme, DNA e proteine formano la cromatina. Gli istoni sono estremamente abbondanti, con decine di milioni di molecole per cellula, per una massa complessiva paragonabile a quella del DNA. Le proteine non istoniche comprendono fattori di rimodellamento della cromatina, regolatori della trascrizione, topoisomerasi, coesine e condensine, tra le altre.
Il primo livello di organizzazione è il nucleosoma, che riduce la lunghezza lineare del DNA di circa un terzo. Al microscopio elettronico, fibre di cromatina isolata mostrano insiemi di particelle nucleoproteiche ravvicinate (Figura 02.03-12). Quando tali fibre vengono parzialmente distese, emerge una successione regolare di unità globulari intervallate da tratti di DNA, evidenziando un’architettura ripetitiva (Figura 02.03-12). Ogni unità centrale, la particella nucleosomica, è composta da un ottamero istonico formato da due copie ciascuna di H2A, H2B, H3 e H4, attorno al quale si avvolgono 147 coppie di basi di DNA a doppio filamento (Figura 02.03-13). Strutture ad alta risoluzione mostrano che il DNA compie circa 1,7 giri sinistrorsi attorno al disco proteico (Figura 02.03-14). Il DNA che collega due particelle successive, detto DNA di connessione (linker), è variabile e può estendersi da pochi nucleotidi fino a circa 80 nucleotidi.
Gli istoni del core sono proteine relativamente piccole, ricche in residui basici (lisina e arginina) che interagiscono con la dorsale zucchero-fosfato del DNA, carica negativamente. Le estremità N-terminali di H2A, H2B, H3 e H4 formano “code” flessibili che sporgono dalla particella nucleosomica e sono bersaglio di numerose modifiche post-traduzionali reversibili (per esempio acetilazione, metilazione, fosforilazione e ubiquitinazione), cruciali per modulare la compattazione della cromatina e il reclutamento di complessi regolatori. L’elevatissima conservazione evolutiva degli istoni core, con minime differenze di sequenza tra specie anche distanti, testimonia il loro ruolo essenziale nella biologia dei cromosomi. Varianti istoniche, come H3.3 o H2A.Z, conferiscono proprietà locali specifiche influenzando ricombinazione, trascrizione e stabilità nucleosomica.
Image Gallery
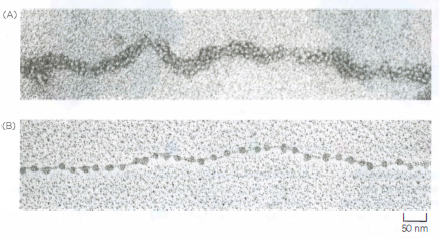
Nucleosomi
I nucleosomi sono visibili al microscopio elettronico. (A) Cromatina isolata direttamente da un nucleo interfascico, che si presenta al microscopio elettronico come una fibra cromatinica composta da nucleosomi impacchettati. (B) Un’altra microfotografia elettronica mostra un tratto di cromatina distesa, decondensata artificialmente e isolata per mettere in evidenza l’aspetto a “collana di perle” dei nucleosomi.
Immagine tratta liberamente da Internet. Se viola i tuoi diritti, contattaci.
Image Gallery
Nucleosomi
I nucleosomi sono visibili al microscopio elettronico. (A) Cromatina isolata direttamente da un nucleo interfascico, che si presenta al microscopio elettronico come una fibra cromatinica composta da nucleosomi impacchettati. (B) Un’altra microfotografia elettronica mostra un tratto di cromatina distesa, decondensata artificialmente e isolata per mettere in evidenza l’aspetto a “collana di perle” dei nucleosomi.
Immagine tratta liberamente da Internet. Se viola i tuoi diritti, contattaci.
Image Gallery
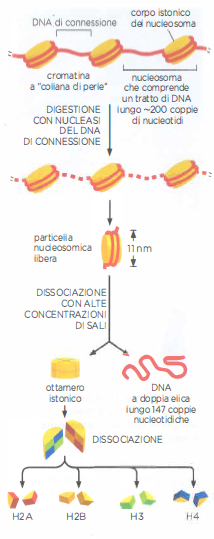
Struttura del nucleosoma
I nucleosomi contengono DNA avvolto intorno a un corpo centrale proteico formato da otto molecole di istoni. In laboratorio la particella nucleosomica si ottiene dalla cromatina digerendola con una nucleasi, un enzima che degrada il DNA in modo da eliminare il DNA esposto, ma non quello avvolto intorno al nucleosoma. Dissociando poi il nucleosoma in DNA e corpo centrale proteico si riesce a determinare la lunghezza del tratto che prima si trovava avvolto: con le sue 147 coppie di basi, il DNA compie quasi due giri intorno al corpo centrale istonico.
Immagine tratta liberamente da Internet. Se viola i tuoi diritti, contattaci.
Image Gallery
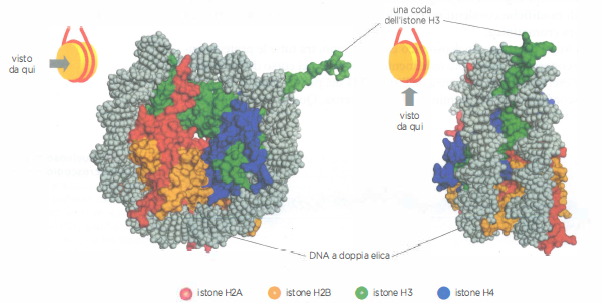
Struttura della particella nucleosomica
La struttura della particella nucleosomica centrale, determinata con l’analisi di diffrazione ai raggi X, mostra come il DNA si avvolga intorno al corpo istonico discoidale. La struttura del nucleosoma è mostrata da due angolazioni diverse. I due filamenti della doppia elica di DNA sono in grigio. Si vede sporgere dal nucleosoma un tratto di coda dell’istone H3 (in verde), mentre le code delle altre proteine istoniche sono state troncate.
Immagine tratta liberamente da Internet. Se viola i tuoi diritti, contattaci.
Il compattamento cromatinico procede per livelli gerarchici
In vivo la cromatina raramente si presenta come una semplice successione lineare di nucleosomi. Le particelle tendono ad associarsi in strutture più addensate, con l’intervento dell’istone H1, detto anche istone di legame, che si connette al DNA di connessione e favorisce l’allineamento ordinato dei nucleosomi, incrementando la compattazione della fibra (Figura 02.03-12). Sebbene l’esatta architettura delle fibre più dense rimanga oggetto di indagine, l’effetto funzionale di H1 e delle code istoniche nel promuovere stati più compatti è ben documentato.
Livelli superiori di organizzazione derivano dalla formazione di anse di cromatina ancorate a una matrice proteica nucleare, con contributi di coesine, condensine e topoisomerasi II; tali anse costituiscono una piattaforma per la regolazione e l’integrazione delle attività genomiche (Figura 02.03-15). Durante l’interfase, l’insieme di anse genera un cromosoma relativamente compatto ma accessibile. All’ingresso in mitosi, ulteriori passaggi di condensazione — inclusa l’azione di condensine che estrudono anse su vasta scala — portano alla tipica morfologia del cromosoma mitotico visibile in metafase (Figura 02.03-16). Questo percorso gerarchico di compattazione consente di alloggiare lunghissime molecole di DNA in volumi nucleari ristretti, preservando al contempo la capacità di attivare rapidamente i processi che richiedono l’accesso al genoma.
Image Gallery
Nucleosomi
I nucleosomi sono visibili al microscopio elettronico. (A) Cromatina isolata direttamente da un nucleo interfascico, che si presenta al microscopio elettronico come una fibra cromatinica composta da nucleosomi impacchettati. (B) Un’altra microfotografia elettronica mostra un tratto di cromatina distesa, decondensata artificialmente e isolata per mettere in evidenza l’aspetto a “collana di perle” dei nucleosomi.
Immagine tratta liberamente da Internet. Se viola i tuoi diritti, contattaci.
Image Gallery
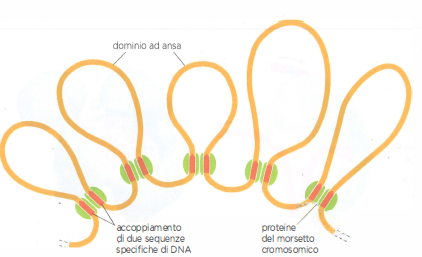
Domini ad ansa della cromatina
Nei cromosomi umani la cromatina è ripiegata in domini ad ansa. Questi domini sono stabiliti da speciali proteine cromosomiche non istoniche che si legano a specifiche sequenze di DNA, creando un morsetto alla base di ciascun anello.
Immagine tratta liberamente da Internet. Se viola i tuoi diritti, contattaci.
Image Gallery
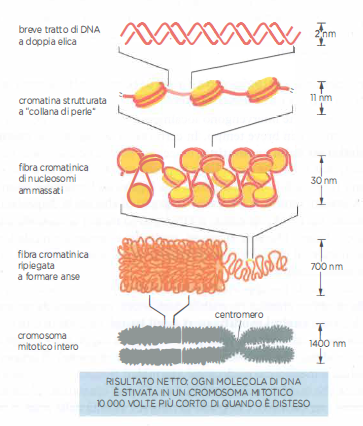
Organizzazione del DNA nel cromosoma
Nei cromosomi il DNA si organizza a parecchi livelli. Lo schema illustra come livelli crescenti di organizzazione del DNA possano dare origine a un cromosoma mitotico altamente condensato. Queste condensazioni sono guidate sia dall’istone H1 sia da un insieme di proteine cromosomiche non istoniche specializzate, tra cui le proteine di bloccaggio dei cromosomi, che formano un’ansa, e la condensina, un’abbondante proteina non istonica. Tuttavia, le strutture reali sono ancora incerte.
Immagine tratta liberamente da Internet. Se viola i tuoi diritti, contattaci.






