


Nei processi di replicazione e riparazione del DNA cooperano numerose proteine
Definizione

Le cellule procariotiche, nella maggior parte dei casi, sono costituite da un unico compartimento racchiuso dalla membrana plasmatica. Le cellule eucariotiche, al contrario, sono suddivise in numerosi ambienti interni delimitati da membrane, che conferiscono un’elevata specializzazione funzionale. Nelle micrografie elettroniche di sezioni di cellule animali e vegetali si distinguono molteplici strutture membranose, di morfologia sacciforme, tubulare, sferica o irregolare, apparentemente distribuite senza una trama geometrica precisa (Figura 05.17-01). Questi compartimenti sono organuli delimitati da membrane, o porzioni degli stessi, ciascuno caratterizzato da un insieme peculiare di macromolecole e metaboliti che ne determinano la funzione. Nei paragrafi seguenti vengono richiamate le principali attività svolte da tali organuli e discusse ipotesi accreditate sulla loro origine evolutiva.
Image Gallery

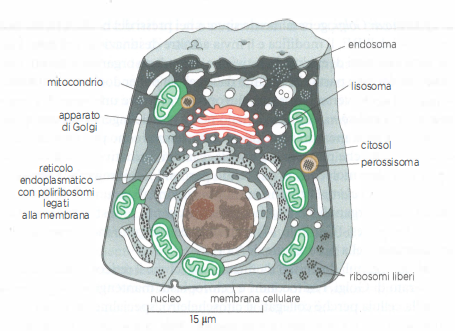
La (Figura 05.17-02) rappresenta i maggiori organuli membranosi di una cellula animale, mentre la (Tabella 05.17-01) ne sintetizza le funzioni principali. Tutti gli organuli sono immersi nel citosol, a sua volta circoscritto dalla membrana plasmatica. Il nucleo, tipicamente l’elemento più evidente, è racchiuso da un involucro nucleare a doppia membrana e comunica con il citosol tramite i complessi del poro nucleare. La membrana esterna dell’involucro è continua con quella del reticolo endoplasmatico (RE), un reticolo di cisterne e tubuli interconnessi che può estendersi in gran parte della cellula ed è il sito principale di biogenesi delle membrane cellulari.
Porzioni estese di RE presentano ribosomi sulla faccia citosolica e prendono il nome di reticolo endoplasmatico ruvido; i ribosomi sintetizzano proteine destinate al lume del RE, alla sua membrana o a organuli dell’endomembrana e alla secrezione, mediante il translocone Sec61. Il reticolo endoplasmatico liscio, privo di ribosomi, è variabile per estensione e svolge funzioni specifiche: sintetizza la maggior parte dei lipidi di membrana, tra cui fosfolipidi, colesterolo e, in tessuti specializzati, ormoni steroidei; contribuisce ai processi di detossificazione, come avviene negli epatociti con sistemi ossidativi a citocromo P450; funge da serbatoio per il Ca²⁺ e ne regola il rilascio e il reuptake tramite canali e pompe dedicate, in stretta relazione con processi come la contrazione muscolare e la trasduzione del segnale.
L’apparato di Golgi, tipicamente localizzato in prossimità del nucleo e del centrosoma, riceve lipidi e proteine dal RE, li elabora (glicosilazione, solfatazione, maturazione dei glicani) e li smista verso la membrana plasmatica, i lisosomi e altri compartimenti. Il Golgi è organizzato in cisterna cis, mediali e trans, con un network trans-Golgi deputato alla selezione e all’impacchettamento dei carichi. I lisosomi sono compartimenti acidi contenenti idrolasi che degradano macromolecole, particelle endocitate e organuli danneggiati; il materiale destinato ai lisosomi transita attraverso endosomi precoci e tardivi, che regolano smistamento e riciclo di recettori e componenti della membrana plasmatica. I perossisomi, piccoli ma metabolicamente attivi, ospitano reazioni ossidative, come l’ossidazione di acidi grassi a catena molto lunga e la sintesi di plasmalogeni; producono perossido di idrogeno, prontamente neutralizzato da catalasi. Mitocondri e, nelle piante e in molte alghe, cloroplasti, sono circondati da doppia membrana e sono specializzati rispettivamente nella fosforilazione ossidativa e nella fotosintesi, grazie a sistemi di membrane interne altamente specializzati per la produzione di ATP.
La posizione di molti organuli, tra cui RE, apparato di Golgi, mitocondri e cloroplasti, è mantenuta tramite ancoraggi al citoscheletro. I microtubuli, insieme ad actina e filamenti intermedi, costituiscono “binari” per il trasporto direzionale di vescicole e organuli, guidato da proteine motorie (kinesine e dineine sui microtubuli; miosine sui filamenti di actina) che utilizzano l’energia liberata dall’idrolisi di ATP. Le interazioni con il citoscheletro coordinano anche la biogenesi e il traffico vescicolare tra gli organuli dell’endomembrana.
Complessivamente, gli organuli membranosi occupano in media quasi metà del volume cellulare e la superficie totale delle loro membrane supera ampiamente quella della membrana plasmatica (Tabella 05.17-02). In una cellula di mammifero, ad esempio, l’area del RE può eccedere di 20–30 volte l’area della membrana plasmatica, rendendo quest’ultima, per estensione e massa, una frazione minoritaria del sistema membranoso complessivo.
La separazione e lo studio funzionale degli organuli richiedono tecniche di frazionamento cellulare. Poiché gli organuli sono troppo piccoli per una manipolazione diretta, si utilizzano metodiche di omogeneizzazione seguite da centrifugazione differenziale e su gradiente di densità. Questo consente di ottenere frazioni arricchite in specifici compartimenti, verificate con enzimi “marker” (per esempio: succinato deidrogenasi per i mitocondri, catalasi per i perossisomi, fosfatasi acida per i lisosomi). Una volta isolati, gli organuli possono essere analizzati per composizione proteica e lipidica e, in condizioni controllate in vitro, per attività funzionali. Ad esempio, mitocondri purificati generano ATP ossidando substrati come il piruvato a CO₂ in presenza di ADP, fosfato inorganico e O₂, mentre vescicole derivate dal RE possono essere impiegate per studiare la traslocazione co-traduzionale o la glicosilazione iniziale delle proteine.
Una proprietà topologica cruciale dell’endomembrana è l’equivalenza tra il lume di RE, Golgi, endosomi, lisosomi e l’ambiente extracellulare: il foglietto luminale corrisponde al foglietto esterno della membrana plasmatica. Tale continuità topologica spiega l’orientamento costante delle proteine di membrana lungo la via secretoria e l’esposizione finale dei domini luminali verso l’esterno della cellula.
Dal punto di vista fisico, l’aumento delle dimensioni cellulari richiede membrane interne per sostenere i flussi di materia ed energia. In un modello semplificato sferico, il rapporto superficie/volume decresce con il raggio, secondo: \[ \frac{S}{V}=\frac{4\pi R^2}{\frac{4}{3}\pi R^3}=\frac{3}{R}. \] Un incremento di \(R\) riduce quindi \(\frac{S}{V}\), ponendo un limite funzionale superato nelle cellule eucariotiche tramite l’espansione delle superfici interne:
- RE ruvido: sintesi e inizio di maturazione delle proteine secretorie e di membrana; biosintesi di gran parte dei lipidi;
- RE liscio: sintesi lipidica e steroidea, detossificazione, omeostasi del Ca²⁺;
- Apparato di Golgi: modificazione post-traduzionale di proteine e lipidi, smistamento e confezionamento del carico;
- Endosomi e lisosomi: internalizzazione, riciclo, degradazione dipendente da idrolasi acide;
- Perossisomi: reazioni ossidative e metabolismo lipidico specializzato, con generazione e detossificazione di H₂O₂;
- Mitocondri e cloroplasti: produzione di ATP tramite fosforilazione ossidativa o fotosintesi.
Image Gallery
| Compartimento | Funzione principale |
|---|---|
| Nucleo | Contiene il genoma e dirige la sintesi di DNA e RNA |
| Citoplasma (citosol) | Sede di numerose vie metaboliche, sintesi proteica e organizzazione del citoscheletro |
| Reticolo endoplasmatico (RE) | Produzione di lipidi e sintesi di proteine da destinare a organelli o alla membrana |
| Apparato di Golgi | Modifica e smistamento di proteine e lipidi per la secrezione o per altri compartimenti |
| Mitocondri | Generazione di ATP tramite fosforilazione ossidativa |
| Lisosomi | Degradazione intracellulare di macromolecole |
| Endosomi | Smistamento del materiale interno dopo endocitosi |
| Perossisomi | Ossidazione di molecole tossiche e metabolismo del perossido di idrogeno |
| Cloroplasti (nelle cellule vegetali) | Sintesi di ATP e fissazione del carbonio durante la fotosintesi |
Funzioni dei compartimenti cellulari delimitati da membrana
Le cellule eucariotiche possiedono organelli specializzati, ciascuno con compiti distinti. La compartimentazione consente la separazione di processi metabolici complessi, garantendo efficienza e controllo. Nucleo, mitocondri e cloroplasti custodiscono sistemi di produzione energetica o di informazione genetica, mentre RE, Golgi, lisosomi e perossisomi svolgono funzioni di sintesi, smistamento e degradazione.
| Compartimento intracellulare | % del volume cellulare | Numero medio per cellula |
|---|---|---|
| Citosol | 54% | 1 |
| Mitocondri | 22% | ~1700 |
| Reticolo endoplasmatico | 12% | 1 |
| Nucleo | 6% | 1 |
| Apparato di Golgi | 3% | 1 |
| Perossisomi | 1% | ~400 |
| Lisosomi | 1% | ~300 |
| Endosomi | 1% | ~200 |
Distribuzione volumetrica degli organelli negli epatociti
Gli epatociti, cellule altamente metaboliche del fegato, presentano una precisa organizzazione volumetrica dei loro organelli. La maggior parte dello spazio è occupata dal citosol, seguito da mitocondri molto numerosi per sostenere l’elevata richiesta energetica. Reticolo endoplasmatico e apparato di Golgi supportano la sintesi e la secrezione proteica, mentre organelli minori come perossisomi, lisosomi ed endosomi contribuiscono a processi di detossificazione, digestione intracellulare e traffico vescicolare.
Le relazioni tra i compartimenti eucariotici moderni risultano più chiare se considerate in chiave evolutiva. I progenitori delle prime cellule eucariotiche erano verosimilmente simili a batteri, provvisti di una sola membrana plasmatica e privi di membrane interne. In tali organismi, come nei batteri attuali, la membrana plasmatica svolgeva attività oggi ripartite tra diversi organuli: sintesi dei lipidi, trasporto, generazione del gradiente protonico e produzione di ATP. Questa organizzazione è compatibile con le ridotte dimensioni cellulari e l’elevato rapporto superficie/volume dei procarioti. Poiché le cellule eucariotiche hanno volumi tipicamente maggiori di 10³–10⁴ volte rispetto a batteri modello come E. coli, l’espansione delle superfici interne è stata probabilmente un prerequisito per sostenere il metabolismo e i flussi di informazione su scala più ampia.
Le evidenze suggeriscono almeno due vie principali di comparsa degli organuli delimitati da membrana. Da un lato, l’invaginazione e la successiva compartimentazione della membrana plasmatica avrebbero dato origine all’involucro nucleare, al RE, all’apparato di Golgi, agli endosomi, ai lisosomi e ai perossisomi (Figura 05.17-03). Questi compartimenti costituiscono nel loro insieme il sistema delle membrane interne, in cui il traffico vescicolare mantiene la comunicazione e la continuità topologica tra lume e spazio extracellulare. In coerenza con questa origine, gli spazi interni dell’endomembrana vengono “trattati” dalla cellula come se fossero un ambiente equivalente all’esterno.
Dall’altro lato, mitocondri e cloroplasti presentano caratteristiche che rimandano a una origine endosimbiotica. Entrambi possiedono un proprio genoma circolare di piccole dimensioni, apparati di espressione proteica batterio-simili e un doppio involucro membranoso; mostrano inoltre una sorprendente affinità evolutiva con proteobatteri (mitocondri) e cianobatteri (cloroplasti). L’ipotesi più accreditata propone che antenati batterici siano stati inglobati da cellule eucariotiche precoci instaurando una simbiosi stabile (Figura 05.17-04). Nel corso dell’evoluzione, gran parte dei geni endosimbiotici sarebbe stata trasferita al nucleo, con conseguente importazione post-traduzionale della maggioranza delle proteine mitocondriali e cloroplastidiali attraverso complessi di traslocazione specializzati (ad es., TOM/TIM nei mitocondri; TOC/TIC nei cloroplasti). Coerentemente con questa storia, mitocondri e cloroplasti non partecipano al traffico vescicolare dell’endomembrana; mantengono tuttavia scambi intensi di metaboliti e lipidi tramite contatti stretti con altri organuli, in particolare con il RE.
Perossisomi e altri compartimenti “ibridi” forniscono ulteriori indizi sull’evoluzione modulare del sistema endomembranoso. I perossisomi possono originare per gemmazione dal RE e accrescersi tramite importazione di enzimi marcati da segnali PTS1/PTS2 riconosciuti da recettori PEX, oltre che per fissione. Ciò suggerisce una plasticità biogenetica che integra vie di formazione de novo e duplicazione. Nel loro insieme, questi percorsi evolutivi multipli hanno prodotto la complessa architettura compartimentale tipica delle cellule eucariotiche contemporanee.
Oltre al trasporto vescicolare, le cellule eucariotiche sfruttano “membrane contact sites” tra organuli, che consentono trasferimenti diretti di lipidi, ioni e segnali senza fusione di membrane. Tali interfacce, regolate da proteine ponte e proteine di trasferimento lipidico, contribuiscono all’omeostasi di calcio, fosfolipidi e colesterolo, integrando il traffico mediato da vescicole con vie non vescicolari.
Image Gallery
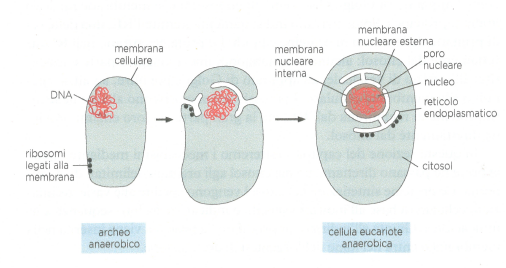
Image Gallery



