


Gli scienziati si avvalgono della bioinformatica per studiare i genomi e le loro funzioni
Definizione

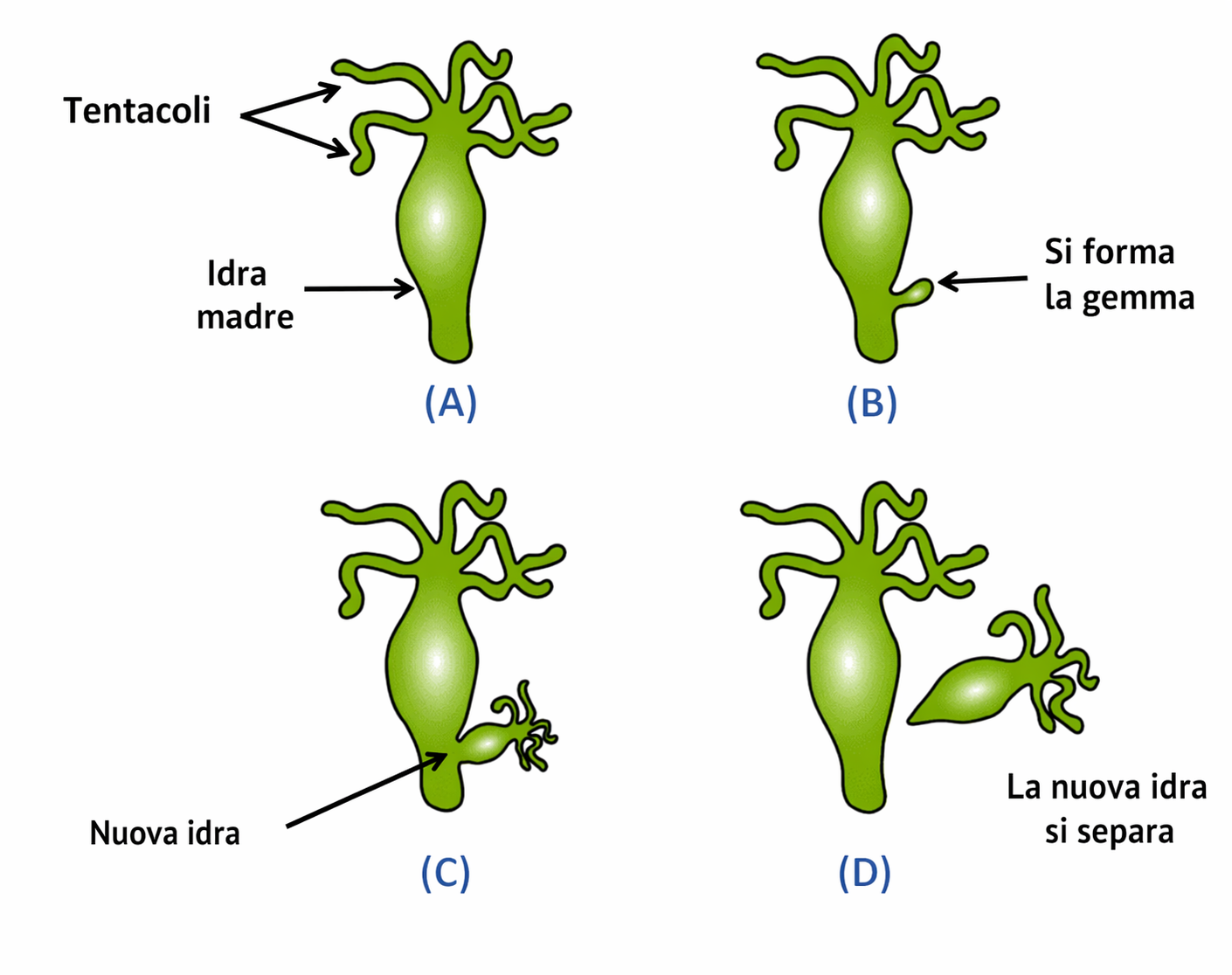
Una larga porzione della biodiversità che ci circonda si propaga attraverso la riproduzione sessuale; tuttavia molte forme di vita, soprattutto microscopiche, adottano strategie asessuate. Nei procarioti e in numerosi eucarioti unicellulari la moltiplicazione avviene per semplice divisione cellulare. Nel mondo vegetale sono diffuse vie asessuate quali stolonizzazione, talea o formazione di bulbi, che generano nuovi individui pluricellulari geneticamente identici alla pianta madre. Anche in animali si osservano modalità asessuate: l’idra, ad esempio, produce nuovi individui per gemmazione (Figura 07.11-01); diversi metazoi, se frammentati, sono in grado di rigenerare le parti mancanti fino a costituire due organismi autonomi. In alcune specie possono inoltre verificarsi scissioni embrionali precoci che danno luogo a individui distinti; tale evento differisce sia dalla fecondazione sia dalla partenogenesi, poiché non prevede l’attivazione di un ovocita non fecondato ma la suddivisione di un embrione già in sviluppo. La riproduzione asessuata è efficiente e conservativa, poiché produce cloni del genitore. La sessualità, al contrario, prevede l’unione di genomi differenti e origina discendenti fra loro non identici e distinti dai genitori. Nei paragrafi seguenti vengono sintetizzati i presupposti cellulari e genetici che rendono possibile tale modalità, adottata dalla grande maggioranza di piante e animali in virtù dei vantaggi evolutivi che ne derivano.
Image Gallery
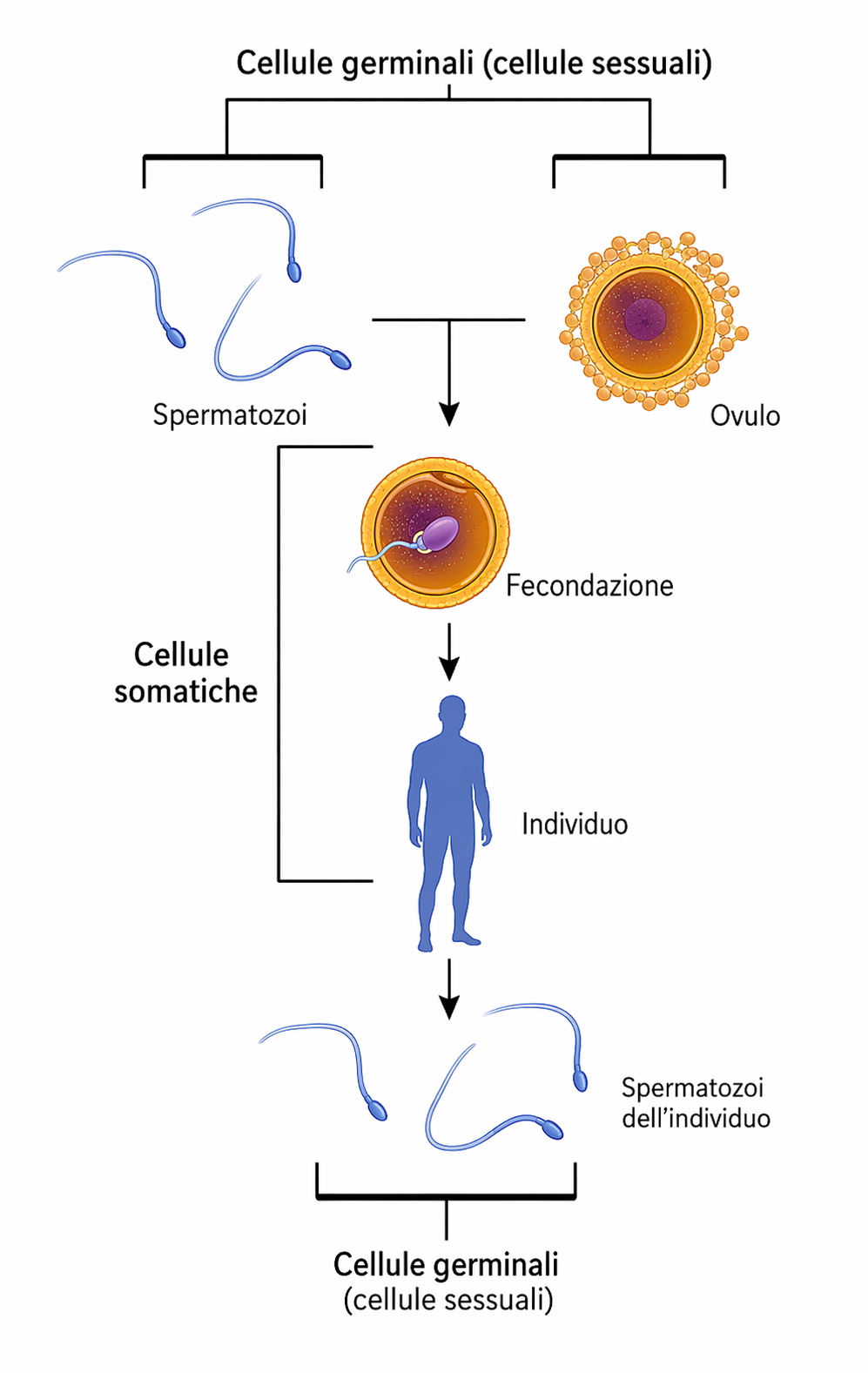
Gli organismi sessuati sono per lo più diploidi: ciascuna cellula somatica possiede due insiemi di cromosomi, uno di origine materna e uno paterna. Le coppie di cromosomi omologhi, ad eccezione dei cromosomi sessuali in specie con determinazione eterogametica, portano lo stesso repertorio genico disposto in loci corrispondenti. Ne consegue che ogni gene è presente in due copie, con l’ovvia eccezione dei geni localizzati su cromosomi sessuali quando sono presenti in singola copia in un sesso. I gameti maturi sono invece aploidi, contenendo un solo assetto cromosomico. La marcata differenziazione dei gameti, detta anisogamia, è tipica degli animali: l’ovocita è voluminoso e privo di motilità, mentre lo spermatozoo è piccolo e dotato di movimento (Figura 07.11-02). La fusione di due gameti aploidi produce lo zigote diploide, che ricompone un nuovo corredo cromosomico a partire dai contributi dei due genitori. Lo zigote avvia quindi una serie di divisioni mitotiche che portano alla formazione di un nuovo individuo con genoma diploide ricombinato (Figura 07.11-03). In molti organismi pluricellulari, incluse le specie di vertebrati, la fase aploide è breve e altamente specializzata alla funzione di vettore genetico. I gameti derivano da precursori diploidi attraverso la meiosi, una divisione cellulare riduttiva che dimezza la ploidia e riorganizza il materiale genetico. La linea di cellule dedicata alla produzione dei gameti è la linea germinale; tutte le altre cellule costituiscono i tessuti somatici e non contribuiscono direttamente alla discendenza (Figura 07.11-04). In altre grandi linee evolutive, come molte piante e alcune alghe, la durata relativa delle fasi aploide e diploide può variare, ma l’alternanza resta il principio cardine del ciclo biologico. Il ciclo sessuale è dunque un’alternanza coordinata tra stati aploide e diploide. Tale alternanza, come discusso di seguito, è alla base della generazione sistematica di novità genotipiche.
Image Gallery

Image Gallery
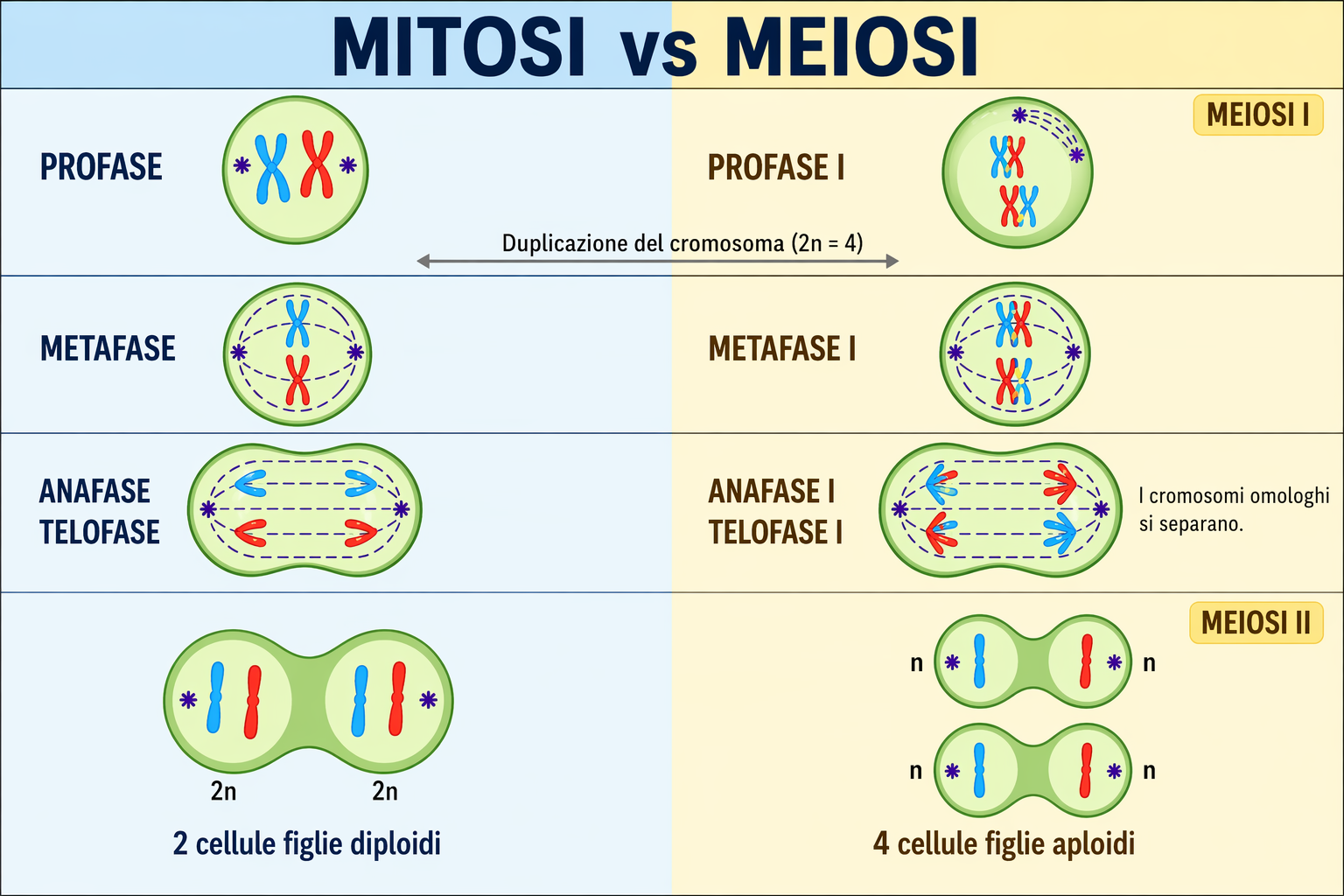
Image Gallery
La sessualità rimescola i cromosomi e gli alleli attraverso eventi meiotici e fecondazione. In meiosi, i cromosomi omologhi di origine materna e paterna vengono separati in modo tale che ciascun gamete riceva una combinazione unica di cromosomi. Poiché i geni su cromosomi omologhi sono gli stessi ma possono presentarsi in varianti diverse, dette alleli, l’assortimento dei cromosomi determina combinazioni alleliche originali nel gamete. La fecondazione, unendo due gameti indipendenti, amplifica ulteriormente la varietà genotipica dello zigote. Due processi fondamentali generano questa diversità:
- assortimento indipendente dei cromosomi omologhi: durante la metafase I meiotica, l’orientamento casuale delle coppie di omologhi sul fuso produce, per il solo assortimento, \(2^n\) combinazioni possibili di cromosomi nei gameti, dove \(n\) è il numero di coppie di cromosomi; per esempio, con \(n=12\) le combinazioni teoriche sono \(2^{12}=4096\);
- ricombinazione omologa (crossing-over): nella profase I i cromatidi omologhi formano chiasmi che consentono scambi reciproci di segmenti, riorganizzando alleli lungo lo stesso cromosoma e generando nuove associazioni intrachromosomiche.
Considerando anche la fecondazione, il numero di combinazioni zigosiche possibili per il solo assortimento sale a \(2^{2n}\); in una specie con \(n=20\), ciò implica oltre un milione di possibili genotipi diversi prima ancora di includere gli scambi ricombinativi. La ricombinazione omologa, la cui frequenza è proporzionale alla distanza genetica tra loci, moltiplica ulteriormente la diversità potenziale spezzando le associazioni alleliche preesistenti. Alle mutazioni de novo, che introducono nuovi alleli a bassa frequenza, la sessualità aggiunge quindi un rimescolamento sistematico su due scale: interchromosomica (assortimento) e intrachromosomica (ricombinazione). Il risultato è un ventaglio di genotipi unici a ogni generazione, sul quale la selezione naturale e altri processi evolutivi possono agire con efficacia.
Poiché i rimescolamenti allelici sono in gran parte casuali, le nuove combinazioni possono talora ridurre l’idoneità, altre volte incrementarla. Perché, allora, la sessualità risulta così diffusa? Le spiegazioni principali, supportate da osservazioni comparative e da modelli teorici, richiamano i benefici del rinnovamento genotipico in contesti ecologici variabili:
- ipotesi della “Regina Rossa”: in presenza di coevoluzione con patogeni e parassiti, cambiare rapidamente combinazioni alleliche, in particolare nei geni coinvolti nell’immunità, riduce la probabilità che genotipi clonali vengano sistematicamente superati da antagonisti in rapida evoluzione;
- adattamento a eterogeneità ambientale: in ambienti imprevedibili, la produzione di una progenie geneticamente diversificata aumenta la chance che almeno alcuni discendenti siano idonei a nuove condizioni fisiche o biotiche;
- superamento dell’interferenza clonale: in popolazioni asessuate, varianti vantaggiose in individui differenti competono senza poter essere combinate; la ricombinazione sessuale può unire mutazioni favorevoli su uno stesso background genetico accelerandone la diffusione;
- limitazione dell’accumulo di mutazioni deleterie (cricchetto di Muller): la ricombinazione facilita la separazione tra alleli nocivi e benefici, aumentando l’efficacia della selezione purificante nel rimuovere varianti svantaggiose;
- selezione sessuale e scelta del partner: preferenze di accoppiamento possono veicolare alleli associati a elevata idoneità, migliorando la qualità media dei genomi trasmessi alla generazione successiva.
Questi principi trovano riscontro in organismi con cicli flessibili. Alcuni lieviti, in condizioni di scarsità di nutrienti, entrano in meiosi e ricombinano per formare spore; linee con difetti nella ricombinazione o nella meiosi mostrano minore capacità di adattarsi a stress protratti rispetto a linee in grado di alternare modalità sessuate e asessuate. Analoghe strategie si osservano in microalghe unicellulari che attivano fasi sessuate in risposta a variazioni ambientali marcate. Oltre ai benefici immediati in ambienti variabili, la sessualità contribuisce a mantenere un equilibrio dinamico nella popolazione: riduce gli squilibri di associazione tra loci, migliora la risposta alla selezione e rallenta l’accumulo irreversibile di mutazioni dannose. Sotto il profilo cellulare, il fulcro di tali vantaggi è la meiosi, nella quale assortimento e ricombinazione ridisegnano di continuo le architetture geniche su cui agiscono i processi evolutivi.


